week-11-hw-bioproduction
Part A: Cell-Free Protein Synthesis | Cell-Free Reagents
For this part I just added one pixel to the artwork.
what you liked about the project, and what about this collaborative art experiment could be made better for next year.
I think this was a great project, it is still to early for me to say what could have been better.

Part B: Cell-Free Protein Synthesis | Cell-Free Reagents
For this assignment we looked at the reagent composition of a cell-free protein synthesis reaction. The reaction is designed to produce fluorescent proteins without living cells. Instead of growing bacteria, the system uses an E. coli lysate that already contains the molecular machinery needed for transcription and translation.
Role of each component
E. coli lysate
BL21(DE3) Star lysate
The lysate provides the biological machinery needed for cell-free protein synthesis, including ribosomes, tRNAs, translation factors, enzymes, and T7 RNA polymerase. This makes it possible to transcribe the DNA template into mRNA and translate the mRNA into fluorescent protein without using living cells.
DNA template
DNA template
The DNA template contains the genetic instructions for the fluorescent protein. In this reaction, T7 RNA polymerase transcribes the DNA into mRNA, and the ribosomes in the lysate translate the mRNA into protein.
Salts and buffer
Potassium glutamate
Potassium glutamate helps recreate the ionic environment of the bacterial cytoplasm. Potassium ions are important for ribosome function, enzyme activity, and general reaction stability.
Magnesium glutamate
Magnesium is especially important in cell-free reactions because it supports ribosome structure, tRNA binding, enzymatic activity, and nucleotide chemistry. Small changes in magnesium can strongly affect transcription and translation efficiency.
HEPES-KOH pH 7.5
HEPES is a pH buffer. It helps keep the reaction close to physiological pH, which is important because enzymes, ribosomes, and fluorescent proteins are sensitive to changes in pH.
Potassium phosphate monobasic and potassium phosphate dibasic
The phosphate salts help buffer the reaction and contribute to phosphate balance. Using both monobasic and dibasic forms helps define the buffering range and pH of the reaction.
Energy and nucleotide system
Ribose
Ribose provides the sugar backbone needed for nucleotide metabolism. In the longer NMP-ribose-glucose system, it helps the lysate regenerate higher-energy nucleotides from lower-energy precursors.
Glucose
Glucose is used as a long-term energy source. The lysate can metabolize glucose to regenerate ATP and support protein synthesis over a longer incubation.
AMP, CMP, GMP, and UMP
These are nucleoside monophosphates. Instead of directly supplying all nucleoside triphosphates, the system can use the lysate’s metabolism to convert these lower-energy nucleotide precursors into the NTPs needed for transcription and energy metabolism.
Guanine
Guanine is a nucleobase that can enter nucleotide salvage pathways. It can be converted into guanine nucleotides, supporting the pool of GTP (guanosine triphosphate) needed for transcription.
Translation mix
17 amino acid mix
The amino acid mix provides most of the amino acid building blocks needed to synthesize the fluorescent proteins.
Tyrosine
Tyrosine is supplied separately because it has low solubility at neutral pH. Supplying it separately makes it easier to control its concentration in the reaction.
Cysteine
Cysteine is also supplied separately because it is chemically sensitive and can oxidize. It is still required as a building block for protein synthesis.
Additive
Nicotinamide
Nicotinamide supports cofactor metabolism, especially NAD-related pathways. This can help maintain metabolic activity in longer cell-free reactions.
Backfill
Nuclease-free water
Nuclease-free water is used to bring the reaction to the correct final volume without adding enzymes that could degrade DNA or RNA.
Difference between the 1-hour PEP-NTP mix and the 20-hour NMP-ribose-glucose mix
The 1-hour PEP-NTP master mix is designed for fast expression. It directly supplies high-energy molecules such as NTPs and uses PEP as an energy source, so it is useful for short, strong cell-free reactions.
The 20-hour NMP-ribose-glucose master mix is designed more like a long-duration metabolic system. Instead of only adding high-energy NTPs directly, it uses NMPs, ribose, and glucose so that the lysate can regenerate energy and nucleotides over time. This is better suited for the 36-hour fluorescence experiment, because the final signal depends not only on fast protein production but also on folding and chromophore maturation.
Bonus: How can transcription occur if GMP is not included but guanine is?
GMP means guanosine monophosphate. It is related to GTP, but it has fewer phosphate groups:
| Molecule | Full name | Phosphate groups |
|---|---|---|
| GMP | Guanosine monophosphate | 1 |
| GDP | Guanosine diphosphate | 2 |
| GTP | Guanosine triphosphate | 3 |
Transcription requires GTP, not necessarily GMP as an added reagent. If guanine is present, enzymes in the E. coli lysate can use nucleotide salvage pathways to convert guanine into GMP, and then further phosphorylate it to GDP and GTP. In this way, guanine can act as an upstream precursor for the GTP needed by T7 RNA polymerase.
Part C: Planning the Global Experiment | Cell-Free Master Mix Design
For Part C, I looked at fluorescent protein properties that could affect the final readout in a cell-free expression system. In this experiment, fluorescence depends on several steps: transcription, translation, protein folding, chromophore maturation, pH stability, and the optical readout of each color channel.
Fluorescent protein properties relevant to cell-free expression
sfGFP
sfGFP, or superfolder GFP, is useful in cell-free systems because it is designed to fold efficiently. This makes it a good robust green fluorescent protein, especially when folding conditions are not perfect. Because it matures rapidly, it should give a strong signal relatively early compared with slower-maturing proteins.
mRFP1
mRFP1 is a monomeric red fluorescent protein. A useful property for this experiment is that red fluorescent proteins can be slower or less efficient than green fluorescent proteins, so the final signal may depend strongly on maturation time and reaction stability over the full incubation.
mKO2
mKO2 is an orange fluorescent protein. A relevant property is its moderate acid sensitivity, meaning that pH drift during the reaction could reduce the final fluorescence signal. This makes pH buffering especially important.
mTurquoise2
mTurquoise2 is a cyan fluorescent protein. It is useful because cyan fluorescence can be bright and stable, but the readout may be affected by optical setup, excitation/emission overlap, and background fluorescence. It is also less acid-sensitive than many other fluorescent proteins, so it may be more robust to moderate pH changes.
mScarlet-I
mScarlet-I is a bright red fluorescent protein with improved maturation compared with earlier red fluorescent proteins. Because red fluorescent protein output still depends on folding and chromophore maturation, it may benefit from reaction conditions that stay active for the full incubation.
Electra2
Electra2 is a blue fluorescent protein. Blue fluorescent proteins can be more challenging to read because they may be more sensitive to background fluorescence, excitation conditions, and photostability. For this reason, improving total protein yield and maintaining stable reaction conditions could be important for getting a clear signal.
A guide to choosing fluorescent proteins
My master mix design
For the second phase of the experiment, I selected eight wells and designed a small screen around the default optimized cell-free master mix. Since the interface only allowed additive changes, I used the default well as the lowest-concentration control condition.
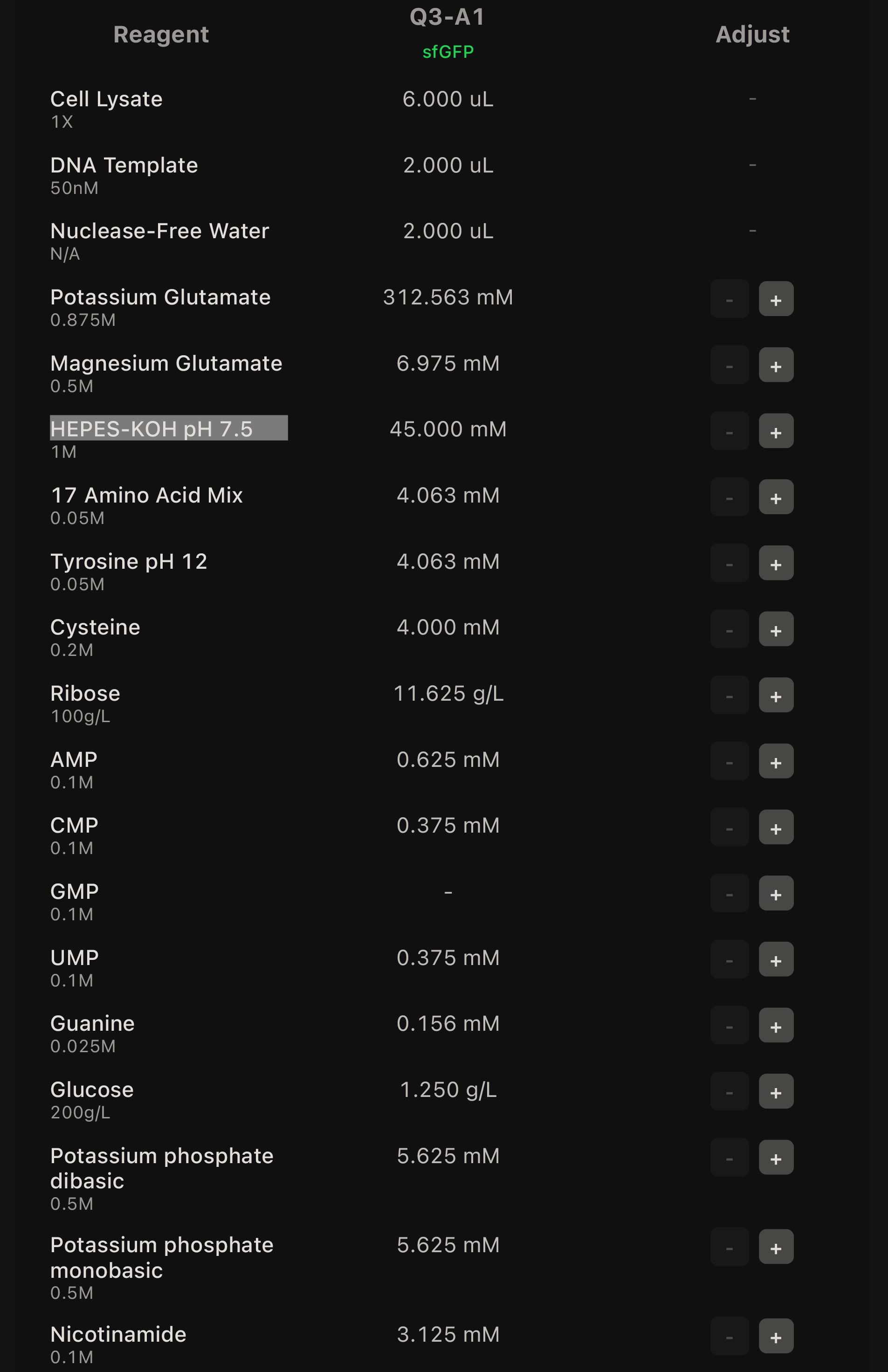
My eight conditions were:
| Well | Condition | Reagent change | Purpose |
|---|---|---|---|
| Q3-A1 | Control | No change | Baseline/default condition |
| Q3-A2 | HEPES high | HEPES-KOH increased from 45.000 mM to 56.250 mM | Tests whether stronger pH buffering improves fluorescence |
| Q3-A3 | Phosphate high | Both phosphate salts increased from 5.625 mM to 7.500 mM | Tests whether extra phosphate buffering/support improves the reaction |
| Q3-A4 | HEPES + phosphate high | HEPES increased to 56.250 mM and both phosphate salts to 7.500 mM | Tests combined pH stabilization |
| Q3-A5 | Magnesium high | Magnesium glutamate increased from 6.975 mM to 8.225 mM | Tests whether higher Mg²⁺ improves transcription/translation |
| Q3-A6 | Magnesium medium | Magnesium glutamate increased from 6.975 mM to 7.600 mM | Tests a smaller Mg²⁺ increase |
| Q3-A8 | Energy regeneration high | Ribose increased from 11.625 g/L to 14.625 g/L; glucose from 1.250 g/L to 1.500 g/L | Tests whether extra metabolic substrate improves long-term expression |
| Q3-A7 | Amino acids high | 17 amino acid mix and tyrosine increased from 4.063 mM to 4.875 mM; cysteine from 4.000 mM to 4.750 mM | Tests whether translation substrates limit protein output |
General considerations for fluorescent protein readout
Fluorescent proteins are useful outputs in cell-free expression systems because they make transcription and translation visible. In BioBits™ Bright, freeze-dried cell-free reactions are used to express fluorescent proteins after rehydration, creating a simple “just-add-water” biological readout. This makes fluorescence a practical way to observe whether the cell-free system is working [1].
However, the strength of the fluorescent signal does not only depend on how much protein is produced. After translation, the fluorescent protein must also fold correctly and form its chromophore before it becomes visibly fluorescent. This means that properties such as folding efficiency, maturation time, brightness, photostability, and excitation/emission wavelength all affect the final signal. Superfolder GFP is an important example because it was engineered to fold more robustly and with improved folding kinetics compared with earlier GFP variants, making it especially useful when reliable fluorescence is needed [2].
Therefore, when choosing a fluorescent protein for a cell-free system, it is important to consider not only the DNA template and expression level, but also the behaviour of the protein after it is made. A protein that folds quickly, matures efficiently, and produces a bright signal will give a clearer and more reliable readout in a paper-based or freeze-dried diagnostic system.
Hypothesis
My hypothesis is that final fluorescence after 36 hours will be improved by reaction conditions that maintain cell-free activity over time, rather than simply increasing early expression. In particular, I expect that improved buffering and sustained energy regeneration may help fluorescent proteins whose final signal depends on folding, pH stability, and chromophore maturation.
I expect the HEPES and phosphate conditions to be useful if pH drift is limiting the fluorescence readout, especially for proteins with acid-sensitive chromophores such as mKO2 or mScarlet-I. I expect the magnesium conditions to affect transcription and translation because Mg²⁺ is important for ribosomes, enzymes, and nucleotide chemistry. However, too much magnesium could also reduce performance by disturbing ionic balance, so the best result may come from the medium magnesium condition rather than the highest one.
I also expect the glucose/ribose condition to support long-duration expression by improving energy and nucleotide regeneration over the 36-hour incubation. The amino acid condition tests whether the reaction becomes limited by translation substrates. If this condition gives higher fluorescence, it would suggest that amino acid availability is limiting protein production.
Overall, my design tests four main variables: pH buffering, magnesium concentration, long-term energy regeneration, and amino acid availability. The goal is to see which type of reagent adjustment best improves final fluorescence in the collaborative cell-free protein artwork experiment.
References
[1] Stark, et. al. (2018). BioBits™ Bright: A fluorescent synthetic biology education kit. Science Advances, 4(8), eaat5107.
https://www.science.org/doi/10.1126/sciadv.aat5107
[2] Pédelacq, et.al (2006). Engineering and characterization of a superfolder green fluorescent protein. Nature Biotechnology, 24, 79–88.
https://www.nature.com/articles/nbt1172