Individual Final Project
1) Abstract
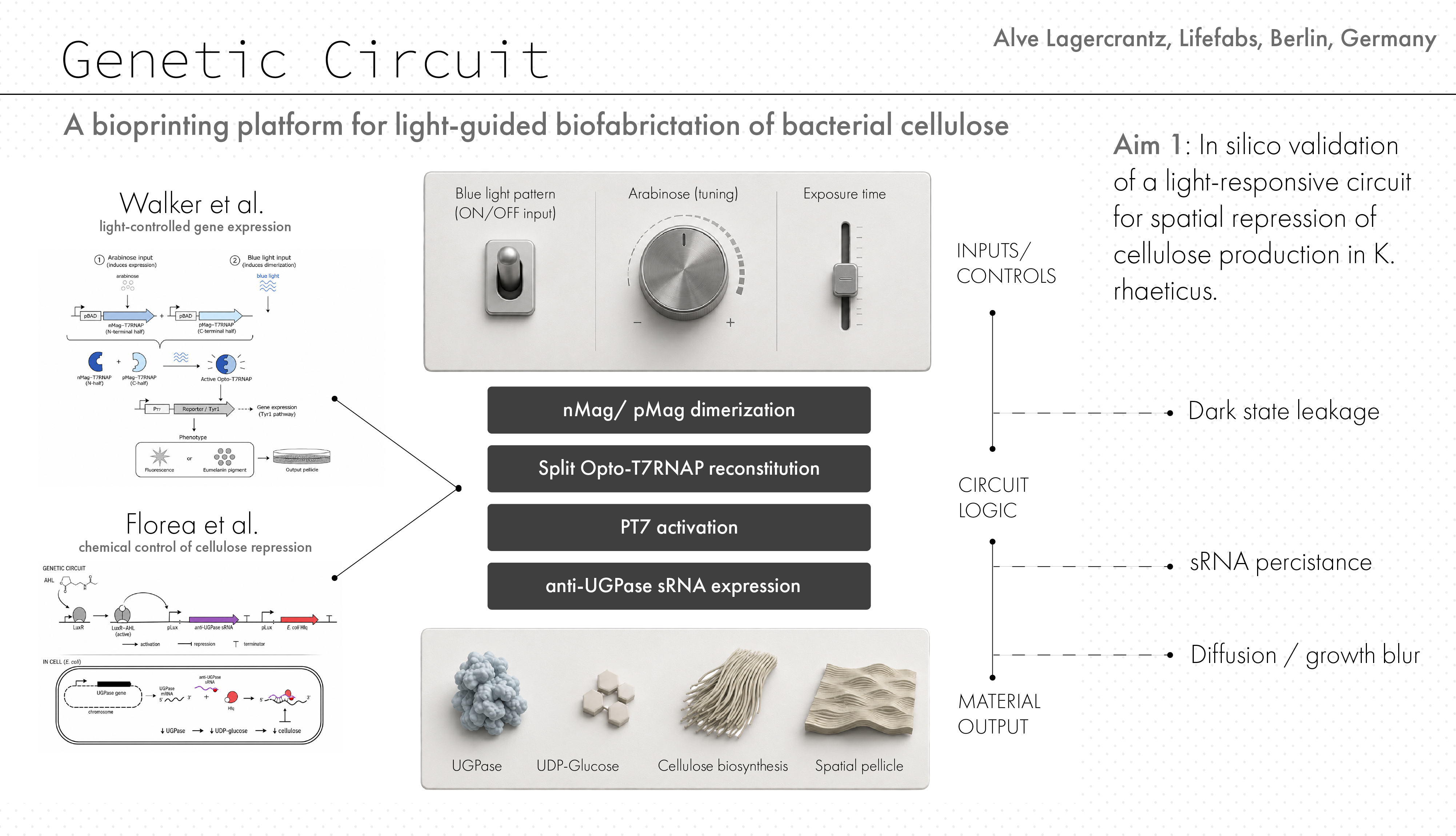
This project explores the use of optogenetic control to encode spatial structure into bacterial cellulose (BC) during growth. Recent advances in engineered Komagataeibacter rhaeticus have demonstrated that gene expression can be patterned within growing pellicles using projected light. However, current approaches primarily focus on pigment production, which is limited in practical textile applications and suffers from low contrast and background expression. This project addresses the need for new fabrication strategies in biomaterials that move beyond surface decoration toward material-level control, which is highly relevant for applications such as textile and bag production.
The overall objective for the project is to develop an optogenetically controlled fabrication platform in which light is used as a programmable input to modulate BC growth and material properties. The central hypothesis is that coupling a light-responsive system to growth-related pathways will enable spatial control over cellulose deposition, resulting in patterned differences in material density, structure, and mechanical behaviour that persist after drying.
To frame the long-term project, I define three aims: first, an in silico validation of the genetic architecture and patterning model; second, future in vitro validation of the circuit; and third, development of a programmable biofabrication platform.
The project will use synthetic biology tools including modular Golden Gate assembly, transformation of K. rhaeticus, and optogenetic control systems. Patterned light exposure will be applied during pellicle growth, and resulting materials will be evaluated in terms of structure, optical appearance, and mechanical behaviour. Together, this work aims to demonstrate a new paradigm for bio fabrication in which light-directed biological growth is used to directly encode function and form into a material.
2) Three Aims
Aim 1: Experimental Aim (In Silico)
To validate, in silico, a light-responsive genetic architecture for spatial control of bacterial cellulose production in Komagataeibacter rhaeticus by combining the Opto-T7RNAP system with an sRNA module targeting UGPase. This aim focuses on establishing the core synthetic biology logic of the project and testing whether projected light patterns could, be translated into localized suppression of cellulose production. The validation for this aim will consist of DNA construct design, circuit architecture, and computational analysis of system behaviour under different conditions of light exposure, arabinose tuning, leakage, and spatial blur. Furthermore, we will develop a construction plan for the hardware development and an experimental plan for the further development of the project in vitro.
Aim 2: Development Aim (in vitro)
The second aim of my final project is to test how the proposed circuit would behave in a growing pellicle. This includes testing, dark-state leakage, sRNA persistence, and pattern diffusion affect the expected spatial resolution and material contrast of the system. As a first step, this system will be tested with a reporter output before being connected to material-regulating genes. Relevant resources include optogenetic systems, the Komagataeibacter Tool Kit (KTK) for modular Golden Gate assembly, and expression control elements from the Acetobacter. The goal is to determine which design parameters are most critical for achieving patterned structural output before wet-lab implementation.
Aim 3: Visionary Aim (in vitro)
The third aim of my final project is to construct a programmable biofabrication platform in which projected light acts as a digital design interface for growing bacterial cellulose materials with encoded structure, density, and interference-based effects such as moiré. This could enable the fabrication of multilayer materials with interference-based effects such as moiré, as well as tuneable optical, tactile, and structural properties. If fully realized, this approach could transform bacterial cellulose from a passive biomaterial into an active, programmable manufacturing system for applications in textiles, fashion, and engineered living materials, where precise control of material properties such as stretch, drape, strength etc is highly desirable. Furthermore such a platform could potentially allow for in vitro “weaving” of fabric which could potentially simplify the complexity of todays supply-chain.
3) Vision, Impact, and Innovation
Vision and impact
The long-term vision of this project is to move bacterial cellulose fabrication from passive sheet growth toward programmable, growth-based manufacturing. If successful, projected light could become a design interface for controlling local material density, thickness, and structure during growth. This could change how biofabricated textiles and soft materials are produced, moving from post-processing and cutting toward in vitro patterning and material formation.
Innovation
The innovative part of this project is not simply using bacterial cellulose as a sustainable material, but connecting optogenetic spatial control to cellulose biosynthesis itself. Existing light-patterned K. rhaeticus systems mainly demonstrate pigment or reporter outputs, while this project proposes material-level control. The project therefore challenges the current model of BC fabrication as uniform pellicle production and reframes it as a programmable, spatially structured manufacturing process.
4) Background and Literature Context
Bacterial cellulose (BC) has increasingly been proposed as a sustainable alternative to petrochemically derived materials in fashion and textile contexts because it can be grown microbially, does not require agricultural land, and offers high purity, crystallinity, tensile strength, biocompatibility, and biodegradability. BC is produced as a dense nanofibrillar pellicle by Komagataeibacter strains and is already being explored across apparel, design, medicine, and electronics [1]. Due to its nanoscale structure, BC is typically fabricated as homogeneous leather-like sheets, although it has also been adapted to more traditional textile processes such as spinning and weaving. However, the limitation of current BC-based textiles is not only material substitution but also manufacturing logic. Existing production systems rely on centralized, large-scale processes. This project addresses that limitation by exploring whether BC can be spatially programmed during growth rather than only modified after production, new materials require new modes of manufacturing.
A key enabling development is the emergence of synthetic biology toolkits for Komagataeibacter rhaeticus. The Komagataeibacter Tool Kit (KTK) provides a modular Golden Gate cloning system for multigene assemblies, while the expanded Acetobacteraceae toolkit provides characterized promoters and regulatory parts for controlled gene expression [2,3]. Together, these systems establish Komagataeibacter as a programmable chassis for engineered living materials.
 [4] Walker, K. T. et al. (2025).
[4] Walker, K. T. et al. (2025).
The most direct precedent for this project is the recent Nature Biotechnology study demonstrating optogenetic patterning in BC-producing bacteria. In this work, K. rhaeticus was engineered to produce melanin through tyrosinase expression, and projected light was used to spatially control gene expression within the growing pellicle [4]. While this study establishes light as a programmable input, the output remains limited to pigment, with issues such as background expression and low contrast. This highlights a key limitation: current systems enable patterned color, but not patterned material structure.
The conceptual foundation for light-based biological patterning originates from Levskaya et al., who demonstrated that projected light can be converted into spatial gene expression patterns in engineered bacteria [5]. This establishes light as a digital interface for biological fabrication.
Parallel work in BC engineering shows that cellulose production itself can be genetically controlled. Florea et al. demonstrated programmable cellulose production and functionalized materials in K. rhaeticus, suggesting that growth and material formation are accessible targets for engineering [6]. However, these approaches have not yet been combined with spatially resolved optogenetic control, leaving a gap in the ability to encode structure during growth.
 moire
moire
The material concept of this project draws on moiré patterns as a structural principle. While moiré effects are widely studied in nanomaterials and optics, particularly in layered systems with offset periodic structures, they have not been explored in BC fabrication. BC’s nanoscale fibrillar network suggests that fine spatial patterning could influence not only visual appearance but also mechanical and structural behaviour [7] much like how different weave structure is changing the material behaviour of fabrics. For this project we call this “light controlled in vitro weaving”.
 SLS/DLP printing
SLS/DLP printing
From a fabrication perspective, this project aligns with developments in additive manufacturing. Techniques such as DLP/SLA printing demonstrate how projected light can define spatial material formation, while architected materials research shows how geometry can control mechanical behaviour. The concept of “Living Manufacture” proposes a microbial fabrication paradigm integrating digital design and biological growth [1].
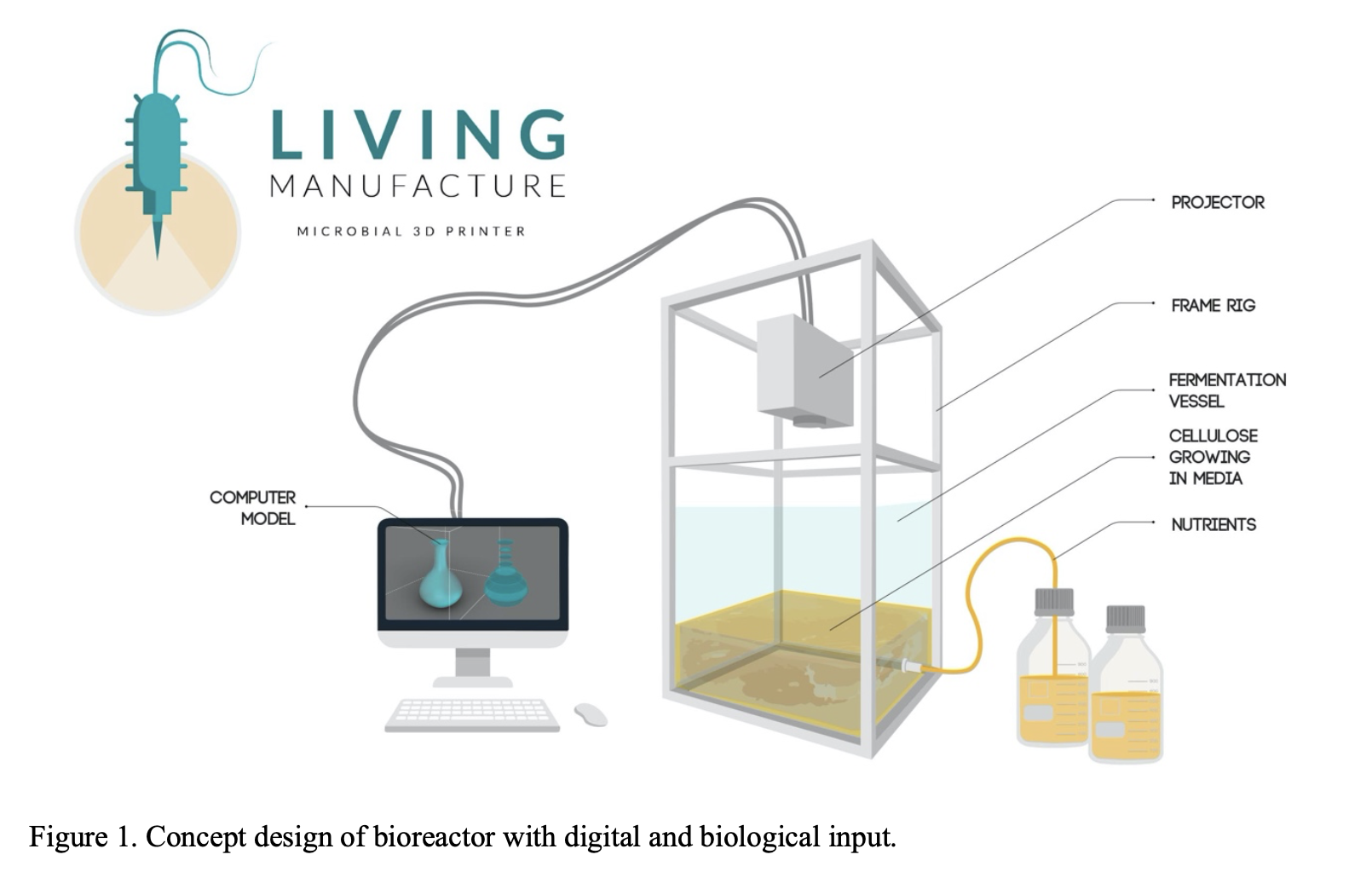 Arnardottir, T. H. et al. Living Manufacture: Principles for a microbial 3D printer.
Arnardottir, T. H. et al. Living Manufacture: Principles for a microbial 3D printer.
Recent advances in bioreactor design further support this approach. One of the main limitations in static bacterial cellulose production is the oxygen starvation of viable cells that occure when the pellicle grow thick leading to an exponential slowdown of pellicle formation.
The mesh dispenser vessel (MDV) bioreactor was developed to circumvent this phenomenon, the design enables top-fed growth, maintaining active cellulose production at the surface and improving layer control [8]. Similarly, the surface airflow (SAF) bioreactor demonstrates that BC material properties can be tuned through process conditions and reactor geometry [9]. This solution would also be interesting for my project since the diffusion of the light through the pellicle is one of the inhibiting factories for controlled high resolution optogenetic prints.
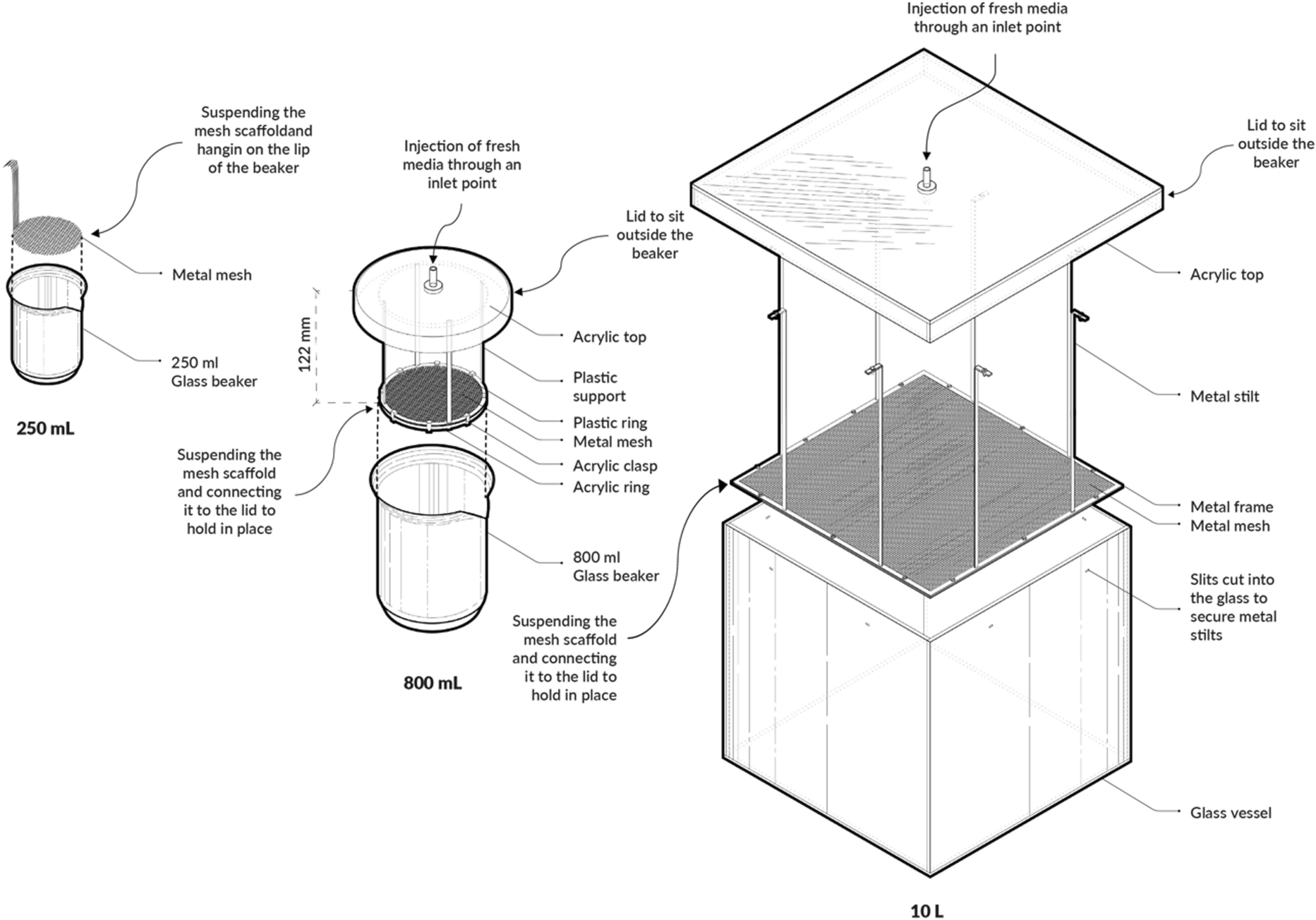 [5] Levskaya, A. et al. (2005).
[5] Levskaya, A. et al. (2005).
Taken together, these studies show that while BC is a promising sustainable material and optogenetic control is feasible, there is currently no system that combines light-based spatial control with growth-driven material structuring. This project addresses this gap by developing a light-directed BC fabrication platform that encodes structure during growth, enabling layered interference effects such as moiré and advancing BC toward programmable biofabrication.
5) Circuit Design, Input and Output Layers
For this project, the optogenetic control system described in [4] is used as a programmable input layer, while the sRNA-based regulation of cellulose biosynthesis from [6] is used as the output layer controlling material formation. By combining these two systems, the aim is to translate spatial light information into spatial control over bacterial cellulose production.
Input
The optogenetic system implemented in [4] is based on a blue-light-sensitive split T7 RNA polymerase (Opto-T7RNAP), originally developed in E. coli and later adapted to Komagataeibacter rhaeticus. A key advantage of this system is that it relies on flavin adenine dinucleotide (FAD) as a chromophore, a metabolic cofactor present in most bacterial cells, including K. rhaeticus. This eliminates the need for additional genetic components for chromophore biosynthesis and makes the system relatively straightforward to transfer across organisms.
Functionally, the T7 RNA polymerase is divided into two inactive fragments (N-terminal and C-terminal), each fused to light-sensitive dimerization domains (nMag and pMag). In the absence of blue light, these fragments remain separate and inactive. Upon illumination at ~450 nm, nMag and pMag heterodimerize, bringing the two halves of the polymerase together and restoring its transcriptional activity. This creates a reversible, light-gated transcription system, where gene expression can be dynamically switched on and off depending on light exposure.
A central feature of this system is the use of the T7 promoter (PT7), which is orthogonal (not interfering with) to the host transcription machinery. Only the reconstituted T7RNAP can recognize and activate PT7, enabling tight and specific control of downstream gene expression. In the original paper, this system was used to drive expression of reporter genes such as GFP or mCherry, as well as functional outputs like tyrosinase (Tyr1) for melanin production.
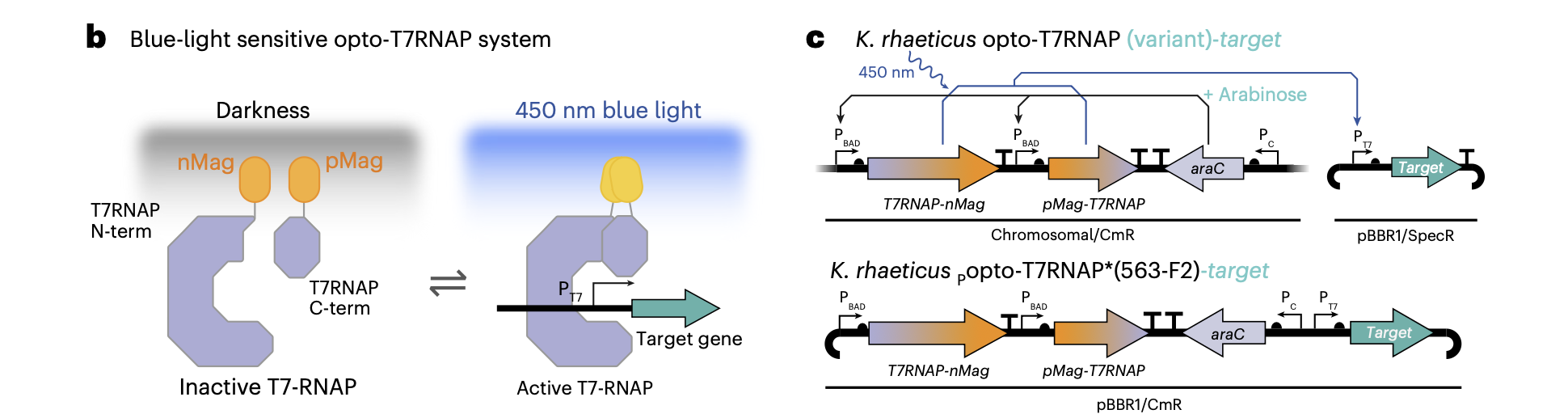 Walker, K. T. et al. (2025)
Walker, K. T. et al. (2025)
Importantly, the system operates with two layers of control. First, arabinose induction regulates expression of the Opto-T7RNAP components via the pBAD promoter. This determines how much of the split polymerase is present in the cell. Second, blue light controls the activity of the polymerase by inducing dimerization (nMag and pMag). In this sense, arabinose does not directly control the output gene but rather tunes the dynamic range of the system. As shown in [4], too high expression levels lead to background activity in the dark (leaky expression), while too low levels reduce overall signal output. Therefore, careful tuning of arabinose concentration is essential to achieve a high contrast between light and dark states.
Footnote:
A dimerization domain is a part of a protein that makes it bind to another protein (or itself)—so that two proteins come together and form a dimer.
- “di-” = two
- “-mer” = unit
Output
For the output layer, the system described in [6] provides a mechanism to directly modulate bacterial cellulose production. In this work, small regulatory RNA (sRNA) is used to target and downregulate UGPase, an enzyme responsible for producing UDP-glucose, the key precursor for cellulose biosynthesis. By reducing UGPase expression, the metabolic flux toward cellulose production is decreased, allowing control over the thickness and density of the resulting material.
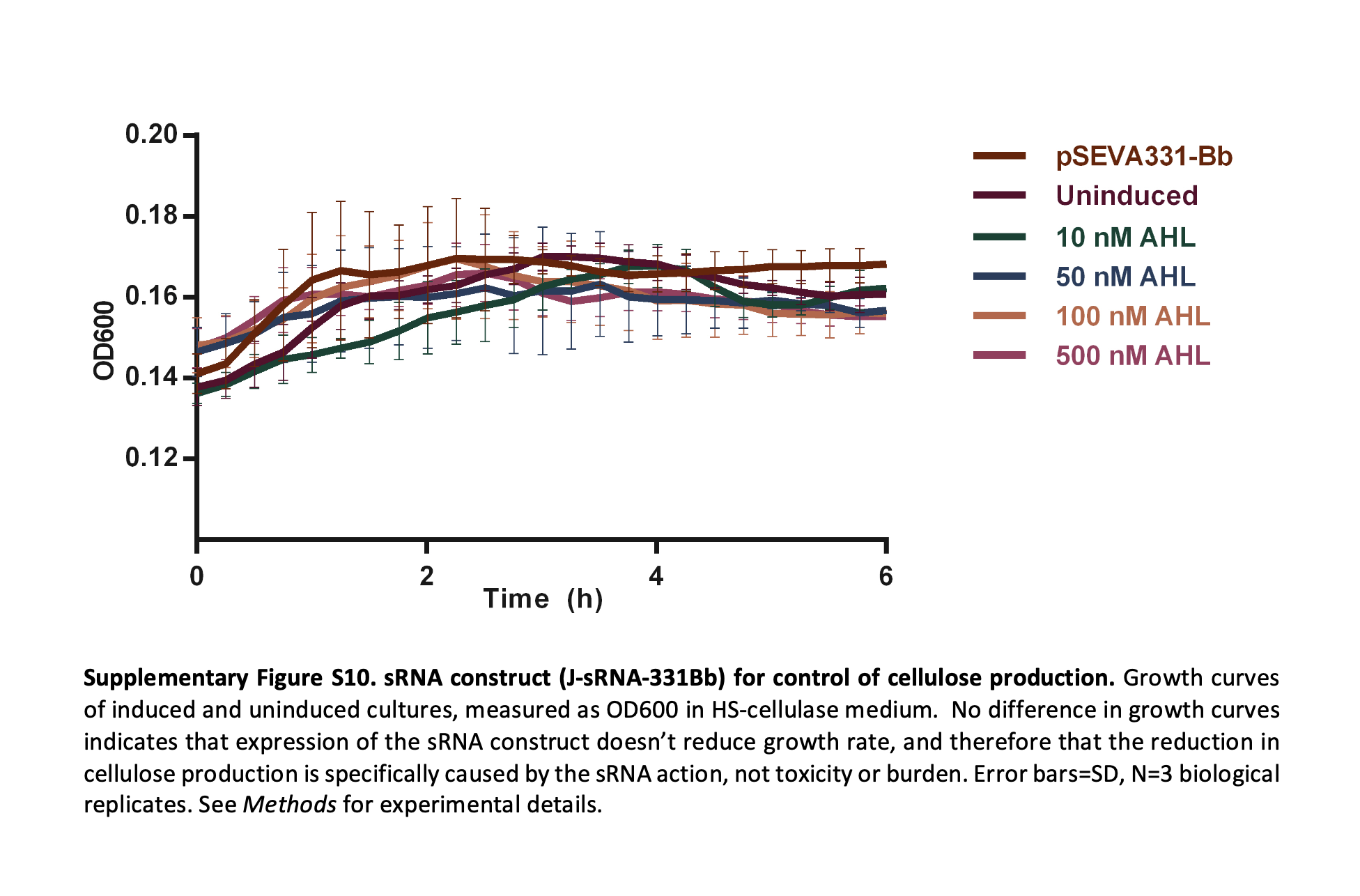 A key insight from [6] is that this intervention does not significantly affect bacterial growth, as shown by unchanged OD600 growth curves across different induction conditions. This decoupling of growth and material production is particularly important for this project, as it enables modulation of material properties without destabilizing the system or reducing viability. Florea, M. et al. (2016).
A key insight from [6] is that this intervention does not significantly affect bacterial growth, as shown by unchanged OD600 growth curves across different induction conditions. This decoupling of growth and material production is particularly important for this project, as it enables modulation of material properties without destabilizing the system or reducing viability. Florea, M. et al. (2016).
In the original implementation, sRNA expression is controlled via a quorum sensing system based on acyl-homoserine lactone (AHL). In this system, AHL binds to the transcriptional regulator LuxR, which in turn activates the pLux promoter to drive sRNA expression. However, for the purposes of this project, this chemical input can be replaced with an optogenetic one by placing the sRNA construct directly under control of the T7 promoter.
This leads to a simplified and more direct architecture:
- From: AHL → LuxR → pLux →sRNA → ↓ UGPase → ↓ cellulose
- To: Light → Opto-T7RNAP → PT7 → sRNA → ↓ UGPase → ↓ cellulose
In this configuration, spatial light patterns directly regulate sRNA expression, which in turn modulates cellulose production locally within the growing pellicle. This avoids the need for intermediate regulatory layers such as LuxR/pLux, reducing system complexity and potential sources of noise or delay.
However, the results from [4] also highlight important limitations that are directly relevant to this project. When using enzymatic outputs such as tyrosinase, the system exhibited significant background activity, leading to melanin production even in the absence of light. This reduced the contrast and resolution of spatial patterning. Similar challenges are likely to arise when controlling cellulose production via sRNA, where leaky expression could lead to global suppression rather than localized modulation.
Achieving high spatial resolution therefore depends on minimizing background expression and maximizing the dynamic range between light and dark states. This can be addressed through several strategies, including tuning arabinose concentrations, optimizing promoter strength, reducing plasmid copy number (e.g., through chromosomal integration), or introducing degradation mechanisms for the sRNA to reduce persistence.
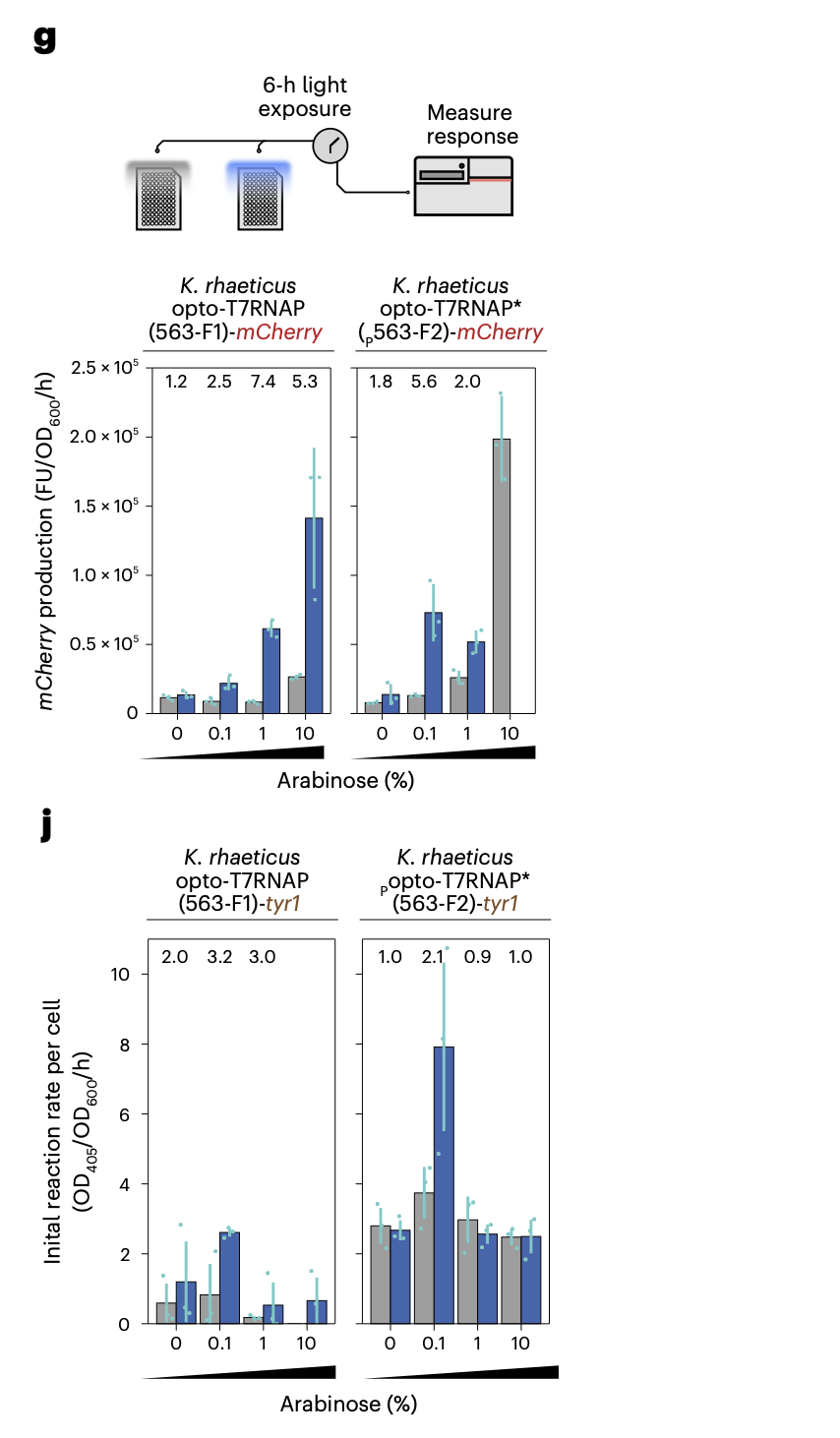
Characterization of optogenetics constructs with mCherry target gene under differing arabinose percentage (wt/vol) concentration. Bars (blue, exposed and gray, unexposed) show mean increase in red fluorescence after 6 h normalized by OD600. Error bars show s.d. of three biological replicates. Fold difference between exposed and unexposed cells is shown above, except in instances of poor growth j, Optogenetic construct characterization with tyr1 under differing arabinose induction. The bars (blue,exposed and gray, unexposed) show the mean and s.d. of three biological replicates of initial (0–100 min) reaction rate of eumelanin production measured at OD405, normalized to the number of initial cells at OD600 at time point 0. source Walker, K. T. et al. (2025).
Note the opto-T7RNAP system demonstrates tuneable, light-responsive gene expression in K. rhaeticus. However, downstream functional outputs (e.g. Tyr1 activity) show nonlinear dependence on induction conditions (see pic above), highlighting a key challenge in translating gene-level control into predictable material outcomes such as bacterial cellulose structure.
Additional constraints arise from the physical properties of the system. In [4], the spatial resolution of patterned gene expression was reported to be on the order of ~0.8 mm, and significant exposure times (up to 40–80 hours) were required to generate visible outputs. These temporal and spatial limitations are particularly relevant for bacterial cellulose, which grows as a continuous pellicle at the air–liquid interface. Over time, diffusion of signalling molecules, enzymes, or metabolic intermediates may further blur spatial boundaries, reducing pattern fidelity.
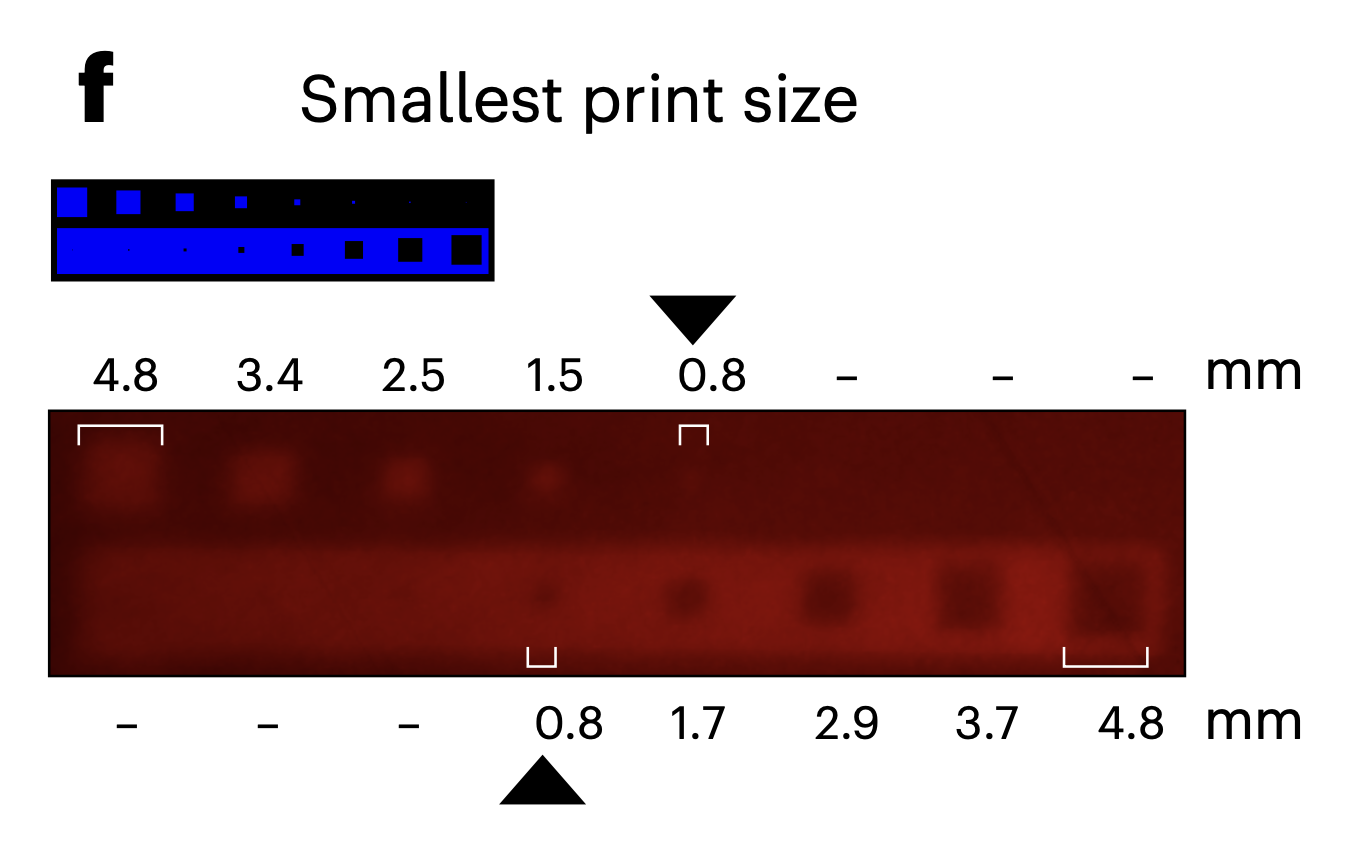 Smallest projected mark on the exposed pellicle. Walker, K. T. et al. (2025)
Smallest projected mark on the exposed pellicle. Walker, K. T. et al. (2025)
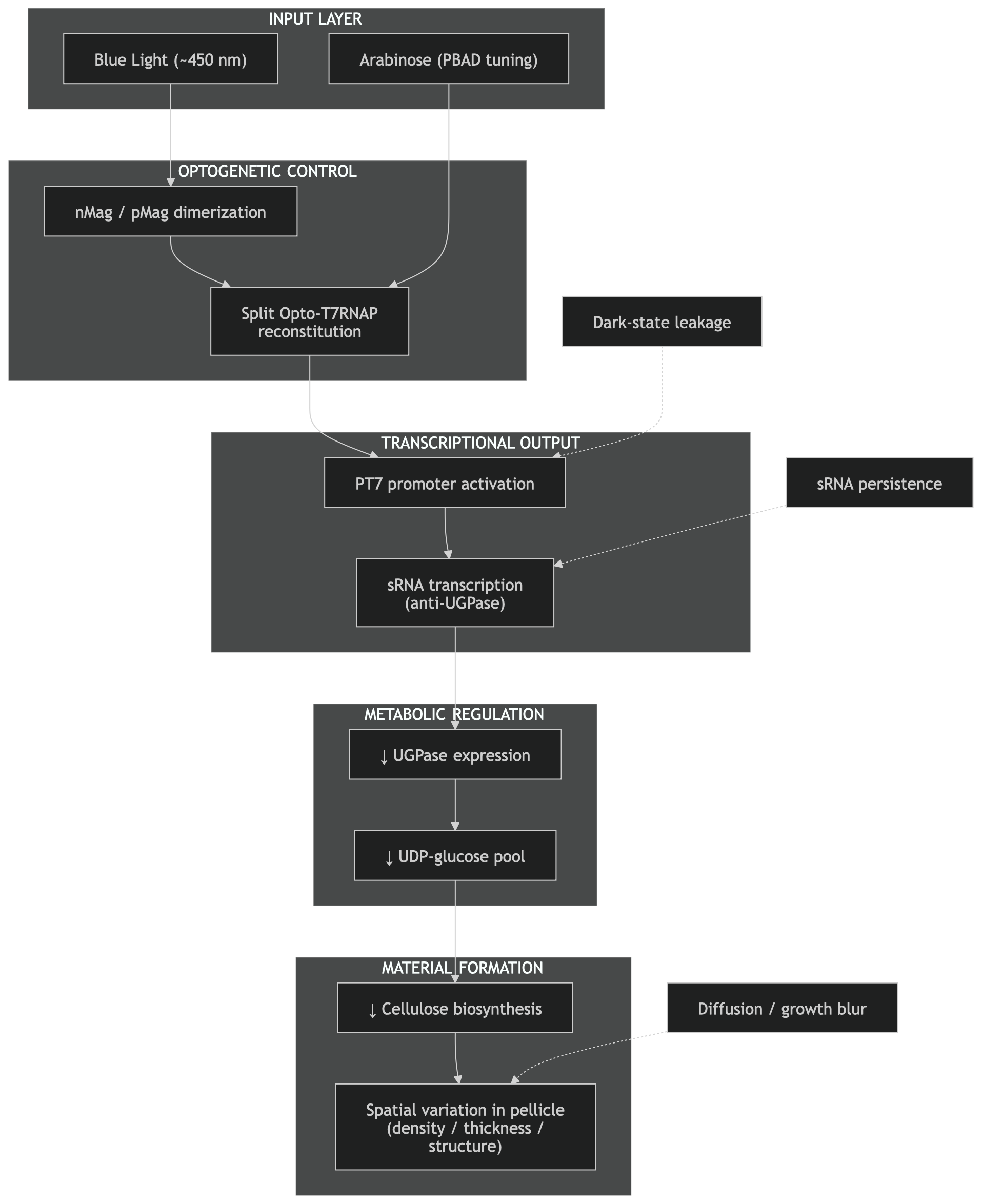 overview of component in my genetic circle (authors own)
overview of component in my genetic circle (authors own)
Taken together, these findings suggest that while optogenetic control of cellulose production is feasible, achieving precise and high-resolution material patterning will require careful optimization at both the genetic and process level. In particular, balancing expression strength, minimizing background activity, and accounting for diffusion and growth dynamics will be critical for translating light patterns into stable material structures.
6) Experimental Design, Techniques, Tools, and Technology
Aim 1 success criteria
- one final circuit architecture
- one DNA design package
- one working simulation Google Colab Notebook
- at least 3 simulated pattern cases
- one quantitative interpretation of leakage / blur limits
- one future wet-lab and hardware development roadmap
Aim 1 validation
- Design the DNA architecture
- Build the construct logic for
Light → Opto-T7RNAP → PT7 → sRNA → ↓ UGPase → ↓ cellulose
- Specify parts, regulatory logic, and construct organization.
- Software: Benchling, Twist, GenBank / NCBI
- Run computational analysis
- Simulate projected light input versus predicted output.
- Based on data from paper [4] and [6] Test how leakage and blur affect pattern contrast.
- Software: Python, Colab notebook, Grasshopper
- Interpret the result biologically
- Show whether the system is likely to support spatial control in principle.
- Identify the parameter window where the design works or fails.
Experimental Plan (in this order)
- finalizing the biological architecture of the proposed system, combining the blue-light-responsive Opto-T7RNAP input layer from Walker et al. with the sRNA-mediated UGPase repression strategy from Florea et al. to create the circuit logic: Light → Opto-T7RNAP → PT7 → sRNA → ↓ UGPase → ↓ cellulose.
- Define the core DNA components required for the system, including the split Opto-T7RNAP modules, the PBAD/arabinose tuning layer, the PT7-controlled sRNA cassette, and the target sequence directed against UGPase mRNA.
- Design the proposed constructs in Benchling
- Translate the biological circuit into a computational model (Colab notebook) that links projected light intensity to predicted transcriptional activation.
- The first layer of the model will represent projected light as a two-dimensional spatial input map, using simple test patterns such as stripes, gradients, dots, and offset line fields relevant to the moiré concept.
- The second layer will model the optogenetic response of Opto-T7RNAP, including both basal dark-state activity and light-induced activity, the literature shows that the system can become leaky depending on arabinose concentration and target gene choice.
- Arabinose will be included as a tuning parameter that changes the total level of Opto-T7RNAP expression and therefore influences both output strength and background activity.
- The third layer of the model will represent sRNA accumulation and persistence over time, using a simplified response function rather than a full kinetic model.
- The fourth layer will convert predicted sRNA levels into relative UGPase repression and then into a predicted cellulose suppression map, representing the expected material-level output of the system.
- Include a spatial blur or diffusion to represent the effects of time integration, biological averaging, and possible spreading of the signal during pellicle growth.
- Test the system under multiple scenarios, including low-, moderate, and high-leak conditions, to determine at which point spatial patterning becomes unreliable.
- I will compare different projected feature sizes in the model to estimate what resolution might be realistically retained once leakage and blur are included.
- I will use these simulations to identify a plausible operating window for the system, including the kinds of patterns most likely to survive and the types of patterns most likely to collapse into uniform output.
- Finally, I will interpret the results as a feasibility analysis for future wet-lab implementation in Komagataeibacter rhaeticus, including a clear decision tree for what should be experimentally tested first if the project is continued after HTGAA.
5. Challenges, limitations, and alternative strategies
- The model does not predict exact biological kinetics.
- Leakage in the real organism may differ from the simulated assumptions.
- Diffusion and pellicle growth dynamics are simplified.
7) DNA Circuit Design
Opto-T7RNAP Cassette Design (Half1 & Half2)
This work focused on constructing two expression cassettes for the split Opto-T7RNAP system in Komagataeibacter rhaeticus, enabling light-controlled gene expression.
Each cassette follows the same modular architecture: AraC + pBAD regulatory module → RBS → OptoT7 fusion protein → rrnB T1 terminator
“Each OptoT7 cassette includes the AraC–pBAD regulatory module, enabling arabinose-inducible expression of the split OptoT7 RNAP halves.”
Sequence sourcing
The AraC/pBAD module was extracted as a complete regulatory unit from a validated plasmid (as described in [4]), rather than rebuilt from individual parts, ensuring correct promoter regulation and arabinose responsiveness. The RBS is a strong ribosome binding site derived from the T7 gene 10 sequence, chosen for efficient translation. The rrnB T1 terminator was selected as a reliable bacterial terminator to ensure proper transcriptional termination.
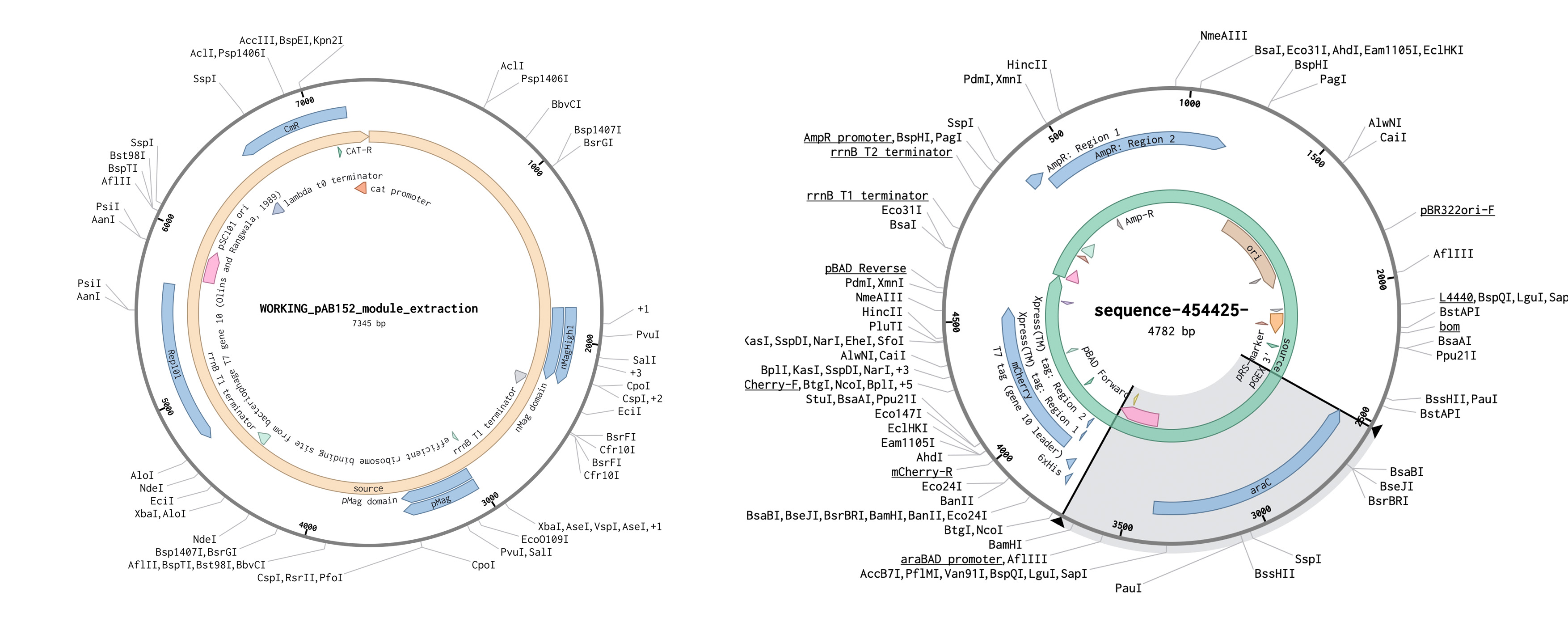 The two plasmids used for the construct
The two plasmids used for the construct
Source1 T7RNAP fragment fused to nMagHigh1 and pMag: https://www.addgene.org/54630/ (same as article [4]) Source2 pBAD: https://www.addgene.org/54630/ picked because it is a well established and tested plasmid
The Opto-T7RNAP split system was implemented as two separate constructs:
- Half1: T7RNAP fragment fused to nMagHigh1
- Half2: T7RNAP fragment fused to pMag
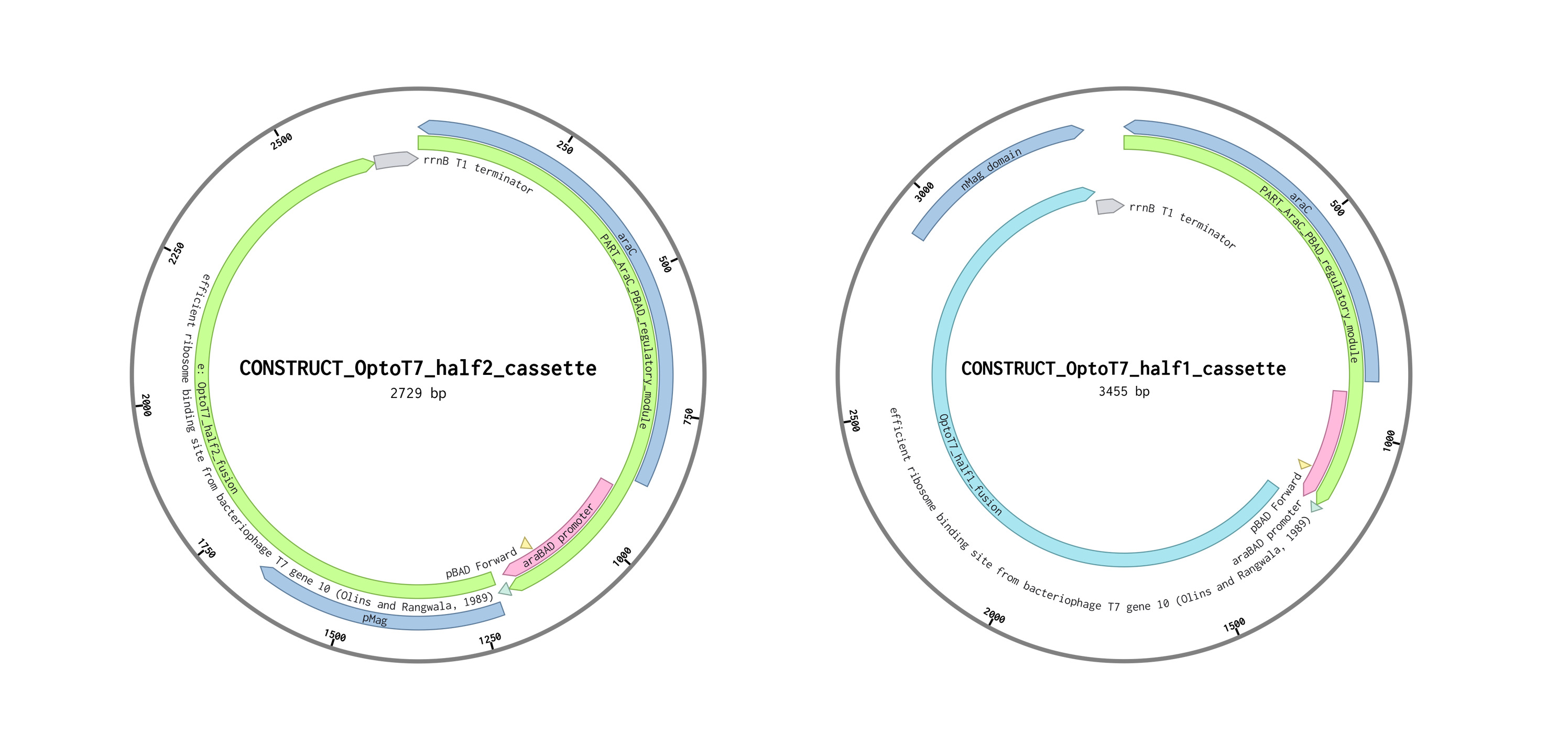 half 1 and 2 Constructs are shown as circular maps for visualization purposes; all sequences were designed and ordered as linear DNA fragments.*
half 1 and 2 Constructs are shown as circular maps for visualization purposes; all sequences were designed and ordered as linear DNA fragments.*
Design strategy
Rather than assembling parts from scratch, the approach was to:
- Extract coherent functional modules from existing, validated constructs preserve native regulatory architecture and minimize risk of mis assembly or unexpected behaviour.
- Both cassettes were finalized as linear, synthesis-ready DNA sequences, with verified structure, correct reading frames, and consistent regulatory logic.
the two cassetes can be found here:
- https://benchling.com/s/seq-IW5krI1Tp60w9H4YLG3P?m=slm-faHhPi0XNjdWFQRu9eTS
- https://benchling.com/s/seq-ozdvCTDIXOeobU2qpBWa?m=slm-TK6zIsI9BJnzYeyE8O1l
Output cassette design rationale
The output layer of this project is adapted from the Florea et al. UGPase silencing construct (J-sRNA-331Bb / BBa_K1321328), which was originally designed to suppress cellulose production in Komagataeibacter rhaeticus using an AHL-inducible sRNA system. In the original construct, LuxR is expressed constitutively, and AHL induction activates expression of both a UGPase-targeting synthetic sRNA and E. coli Hfq. The sRNA contains a 5′ sequence complementary to the UGPase mRNA RBS region and a 3′ Hfq-binding region, allowing Hfq-mediated translational repression of UGPase and thereby reducing cellulose production.
Note *Hfq is an RNA-binding protein that:
- stabilizes small RNAs (sRNAs)
- brings sRNA + target mRNA together
- helps them base-pair efficiently*
For this project, the output mechanism is kept as close as possible to the Florea design, but the input logic is rewired. Instead of using LuxR/pLux/AHL, the output will be placed under Opto-T7RNAP control. This means the final adapted output architecture is:
- PT7 → UGPase-targeting sRNA → terminator
- PT7 → E. coli Hfq → terminator
This preserves the original silencing logic while integrating it into a light-responsive genetic system intended for spatial control of cellulose production.
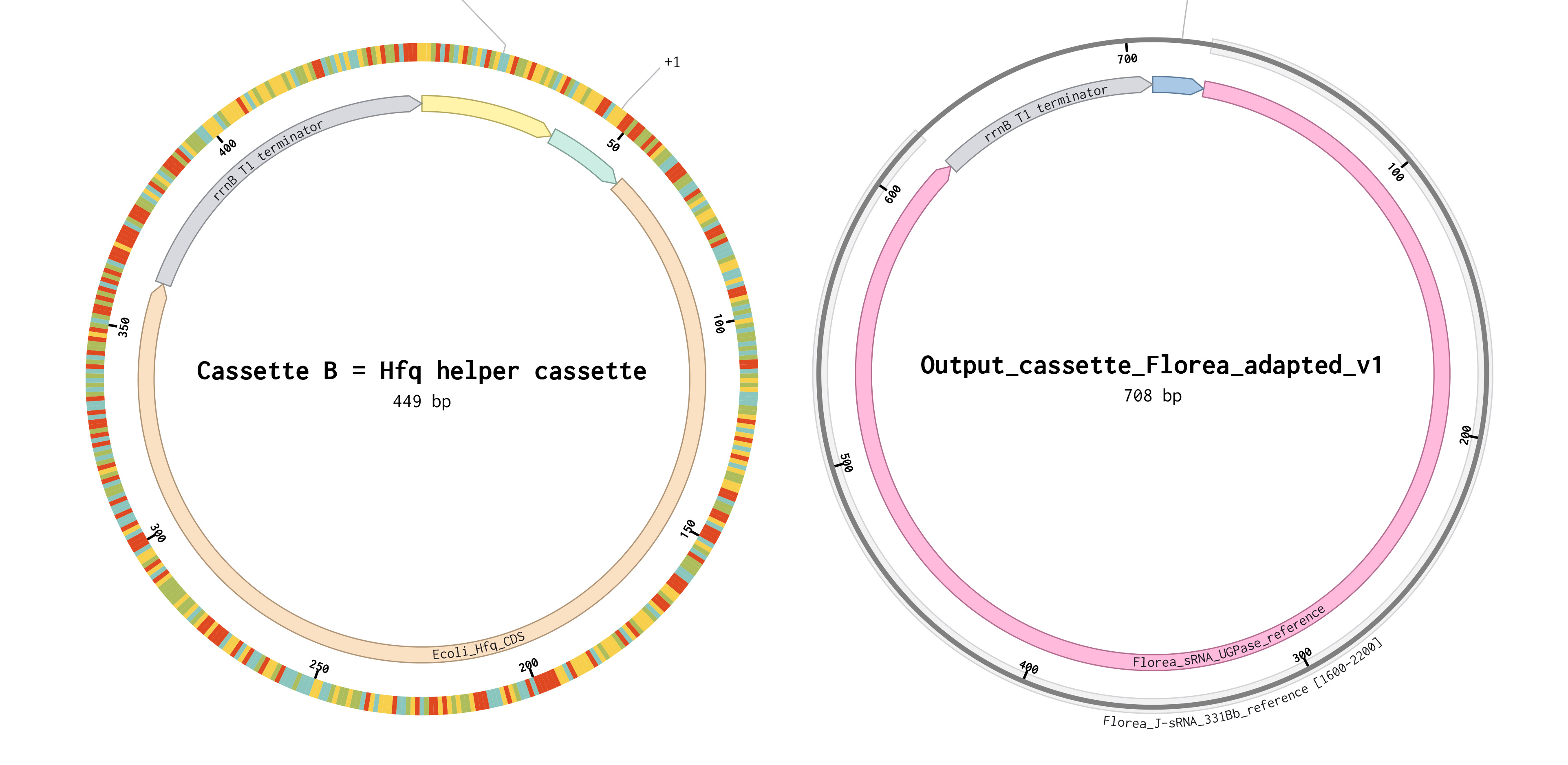 Constructs are shown as circular maps for visualization purposes; all sequences were designed and ordered as linear DNA fragments.
Constructs are shown as circular maps for visualization purposes; all sequences were designed and ordered as linear DNA fragments.
Annotation note
Because the Registry / toolkit sequence is housed in the pSEVA331-Bb (a low-copy, stable plasmid) context and not presented in the paper as a fully base-resolved minimal feature map, the exact minimal nucleotide boundaries of the synthetic sRNA are not directly obvious from the paper figures alone. Therefore, this project reuses the published functional module logic from Florea rather than claiming a fully independently rederived minimal sRNA core from the schematic
So:
- minimal sRNA boundaries were not independently remapped base-by-base
- Hfq helper cassette was added as a separate constitutive module
- design is in silico and not yet experimentally validated
Note:
- SEVA = Standard European Vector Architecture
- Modular plasmid system designed for synthetic biology
Cassete B= Hfq helper cassete: https://benchling.com/s/seq-rCaVP0yYtscWeJatsMno?m=slm-2WFD8BIFwapoau6hUpCz
Output_sRNA_cassette: https://benchling.com/s/seq-84ojC9Fag5k6a36PuKDG?m=slm-M3qlgER1ZUeCJPtLXprw
DNA Synthesis Strategy
All four genetic constructs designed in this project were evaluated for synthesis feasibility using Twist Bioscience as linear double-stranded DNA fragments. The constructs were submitted as individual sequences in FASTA format without plasmid backbones, corresponding to modular expression cassettes.
The following fragments were included:
- OptoT7_half1 AraC/pBAD input cassette
- OptoT7_half2 AraC/pBAD input cassette
- PT7-driven sRNA output cassette targeting UGPase
- Constitutive Hfq helper cassette
All sequences were provided in 5′→3′ orientation and structured according to standard genetic architecture (promoter → functional element → terminator), with the exception of the sRNA cassette, which is a non-coding regulatory construct.
Note: A non-coding regulatory construct is a DNA module that produces RNA which regulates other genes, rather than being translated into a protein.
Synthesis Outcome
All four constructs passed automated synthesis screening and were accepted under standard synthesis conditions. The total estimated cost for synthesis was approximately $613.83, corresponding to four gene fragments ranging from 461 bp to 3455 bp in length. Minor warnings were observed for the sRNA cassette due to the absence of a conventional open reading frame (ORF), as the sequence encodes a functional regulatory RNA rather than a protein. These warnings did not affect synthesis feasibility and are expected for non-coding constructs.
Twist order
Sequence Design Constraints
Commercial DNA synthesis imposes several constraints on sequence composition, which were considered during construct design:
- GC Content Sequences must typically fall within a moderate GC range (~25–65%) to ensure efficient synthesis and amplification. All constructs in this project fall within acceptable GC ranges:
- sRNA cassette: ~43%
- Hfq cassette: ~47%
- OptoT7 constructs: ~50–52%
Repetitive Elements Long or highly identical repeats can lead to recombination or synthesis failure. The OptoT7 constructs contain a shared AraC/pBAD regulatory module (~1.2 kb), which introduces partial sequence redundancy between fragments. While this did not prevent synthesis acceptance, such repetition could pose challenges in downstream cloning or genomic integration and would typically be minimized in large-scale implementations.
Secondary Structure Strong secondary structures (e.g., hairpins in GC-rich regions) can interfere with oligo assembly. No significant secondary structure issues were flagged during synthesis screening, suggesting that all sequences are within acceptable structural complexity limits.
Homopolymer Runs Long stretches of identical nucleotides (e.g., AAAAAA) can reduce synthesis fidelity. No problematic homopolymer regions were identified in any of the constructs.
Coding vs Non-Coding Sequences Automated synthesis platforms often assume protein-coding sequences and may flag constructs lacking clear open reading frames. The PT7-sRNA output cassette triggered such warnings, as it is intentionally non-coding. This highlights a limitation of automated synthesis validation tools when applied to regulatory RNA systems.
Design Considerations and Trade-offs
Several design decisions influenced synthesis feasibility:
- Inclusion of AraC regulatory module, The OptoT7 constructs include the full AraC/pBAD system rather than a minimal promoter. This increases construct size but ensures functional arabinose responsiveness.
- Separate Hfq expression cassette, Hfq was implemented as an independent module to support sRNA function, increasing modularity at the cost of an additional fragment.
- Use of reference sRNA sequenceThe sRNA cassette was derived from a published system and not fully minimized, prioritizing functional reliability over sequence optimization.
Limitations and Future Optimization
While all constructs passed synthesis screening, several optimizations could improve robustness in future iterations:
- Reducing redundancy between OptoT7 constructs to minimize recombination risk
- Codon optimization of coding sequences for Komagataeibacter rhaeticus
- Minimization of non-essential regulatory regions
- Experimental validation of promoter activity and sRNA efficiency in the target organism
Conclusion
The successful synthesis validation of all four constructs demonstrates that the proposed optogenetic system is compatible with current commercial DNA synthesis capabilities. This supports the feasibility of translating the in silico design into a physical genetic system for experimental implementation.
| Fragment | Function | Status |
|---|---|---|
| OptoT7 half 1 | Light-responsive T7RNAP component | Designed |
| OptoT7 half 2 | Light-responsive T7RNAP component | Designed |
| PT7-sRNA | Represses UGPase mRNA | Designed |
| Hfq helper cassette | Supports sRNA function | Designed |
| Full assembled plasmid | Final combined construct | Not built |
| Transformation into K. rhaeticus | Wet-lab validation | Future work |
8) Computation Design
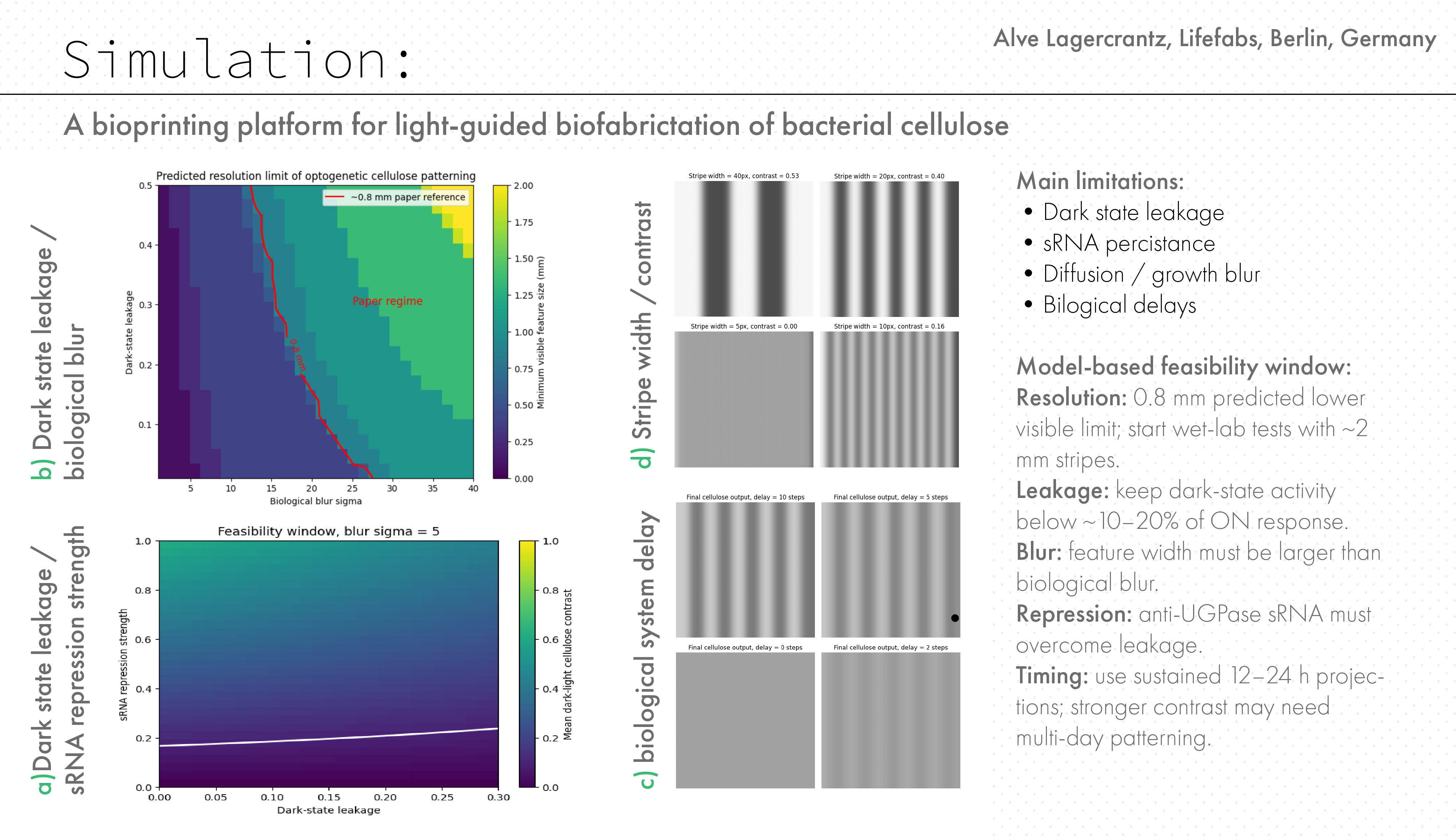
Based on data from Walker, K. T. et al. (2025) initial simulations were performed using periodic stripe patterns of varying feature sizes. The model shows that pattern fidelity strongly depends on feature size.
Simulation of spatial pattern propagation for different feature sizes. Small features (~0.2 mm) are lost due to biological blur, while larger features persist.
As shown in figure above:
- Small features (5 px ≈ 0.2 mm) disappear completely
- Intermediate features (20 px ≈ 0.8 mm) remain partially visible
- Large features (50 px ≈ 2.0 mm) are clearly preserved
Simulation of spatial pattern plotted agains real world data Walker, K. T. et al. (2025). This demonstrates that the system exhibits a minimum resolvable feature size, below which spatial information is lost.
Effect of Biological Parameters
To understand what controls this resolution limit, a parameter sweep was performed across:
- Dark-state leakage (background gene expression)
- Biological blur (representing diffusion, growth, and material integration)
The resulting phase diagram shows the predicted minimum visible feature size (mm) across parameter space. In this model, biological blur sigma is a simplified spatial parameter representing the loss of pattern sharpness between the projected light input and the final cellulose output. It combines possible effects from light scattering, molecular diffusion, cell growth, delayed gene expression, sRNA persistence, and material integration during pellicle formation. The parameter is implemented as the sigma value of a Gaussian filter applied to the predicted cellulose map. It is therefore not a directly measured biological value, but a sensitivity parameter used to estimate how much spatial spreading the system could tolerate before the pattern collapses. Because sigma is defined in pixels, it must be converted to physical units using the model scale of 0.04 mm per pixel. For example, a blur sigma of 20 pixels corresponds to 0.8 mm. This allows the simulated blur range to be compared to the approximately 0.8 mm resolution reported for optogenetic patterning in the Walker et al. system
Predicted resolution limit as a function of dark-state leakage and biological blur. A reference contour (~0.8 mm) was overlaid based on previously reported optogenetic patterning in K. rhaeticus (tyrosinase-based pigment system).The model predicts that this resolution lies along a continuum of parameter combinations, rather than representing a fixed system property.
Key observations: Increasing leakage reduces contrast between patterned regions Increasing blur reduces spatial resolution through mixing The combination of both creates a sharp transition between:
- a regime where patterns are preserved
- a regime where patterns collapse
The code below is included for reproducibility; the main biological interpretation is summarized above each plot.
Code: Stripe resolution simulation
Code: Resolution phase diagram with paper reference
Feasibility Window for Light-Controlled Cellulose Repression
To evaluate whether the proposed optogenetic sRNA system could theoretically generate visible material patterns, I built a simple in silico feasibility model. The goal of the model was not to prove that the biological system will work experimentally, but to estimate under which conditions light-controlled repression of cellulose production could remain spatially visible.
The model simplifies the proposed circuit as a chain of interactions:
light exposure → Opto-T7 activation → sRNA expression → UGPase repression → reduced cellulose production
In this simplified model, illuminated regions produce higher levels of sRNA, which represses UGPase and therefore reduces cellulose production. Dark regions are expected to produce less sRNA and therefore more cellulose. The resulting cellulose pattern should therefore be the inverse of the projected light pattern.
The simulation varies three important parameters:
- dark-state leakage: unwanted sRNA expression in non-illuminated regions
- repression strength: how strongly the sRNA reduces cellulose production
- spatial blur: loss of pattern sharpness caused by light scattering, biological diffusion, growth-layer averaging, or imperfect material response
First, I generated a simple stripe pattern as the projected light input. This was converted into predicted sRNA activity, then into predicted cellulose production. A Gaussian blur was then applied to represent spatial spreading in the biological and material system.
 Input light pattern.
Input light pattern.
A binary stripe pattern was used as the projected light input for the simulation. White regions represent light exposure, while dark regions represent no light exposure.
 Predicted cellulose output after spatial blur.
Predicted cellulose output after spatial blur.
The projected light pattern is converted into predicted cellulose production, where illuminated regions show reduced cellulose production due to sRNA-mediated repression. Gaussian blur was applied to approximate light scattering, diffusion, and biological averaging during pellicle growth.
The model then calculated pattern contrast by comparing cellulose production in intended dark and light regions. This is important because a pattern can only be useful for material structuring if there is a measurable difference between regions of high and low cellulose production.
The resulting heatmaps show a “feasibility window” for the proposed system. The x-axis represents dark-state leakage, and the y-axis represents sRNA repression strength. The colour indicates predicted material contrast, while the white line marks the minimum contrast threshold required for visible patterning.
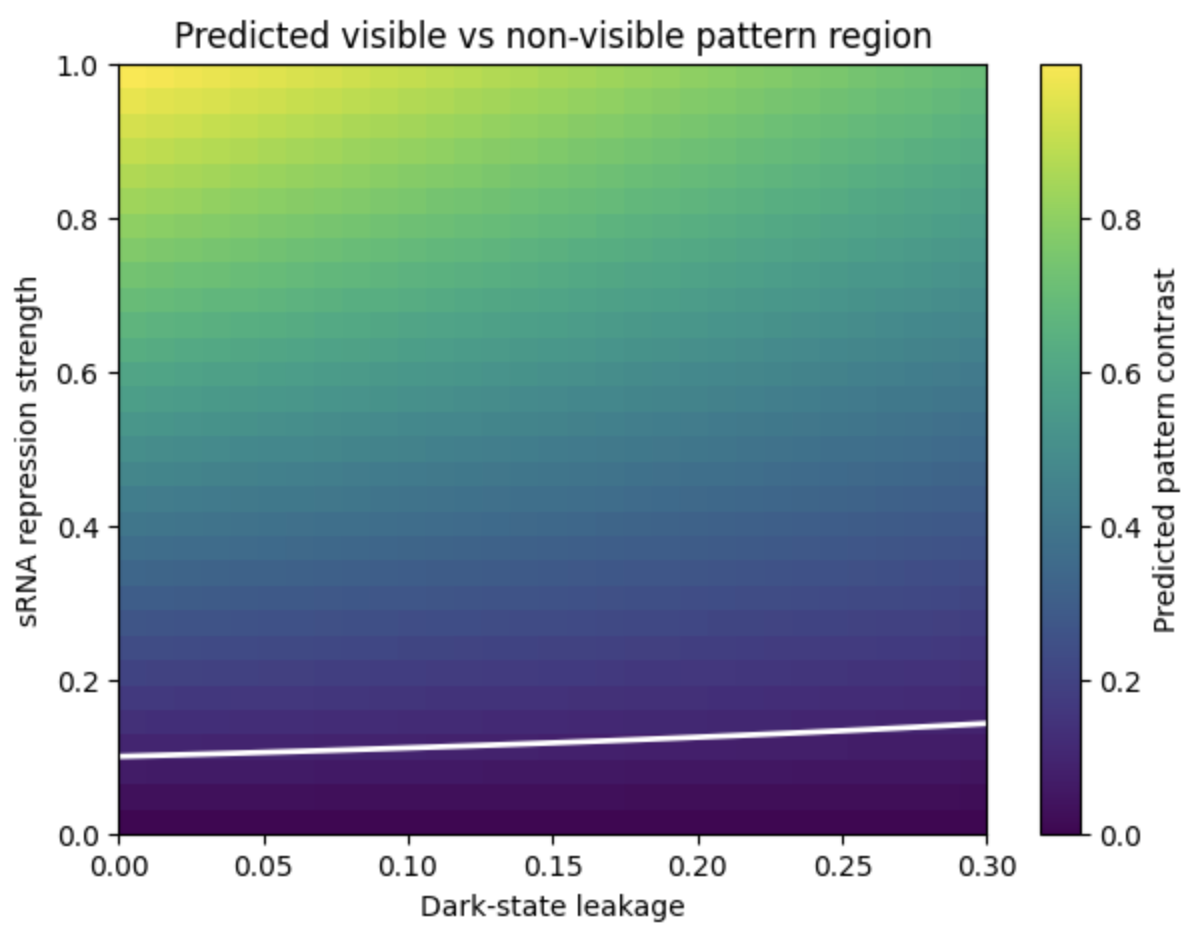 Feasibility window for light-controlled cellulose repression.
Feasibility window for light-controlled cellulose repression.
The heatmap shows predicted cellulose pattern contrast as a function of dark-state leakage and sRNA repression strength. The white contour marks the minimum contrast threshold for visible pattern formation. The model suggests that visible patterning requires low leakage and sufficiently strong repression.
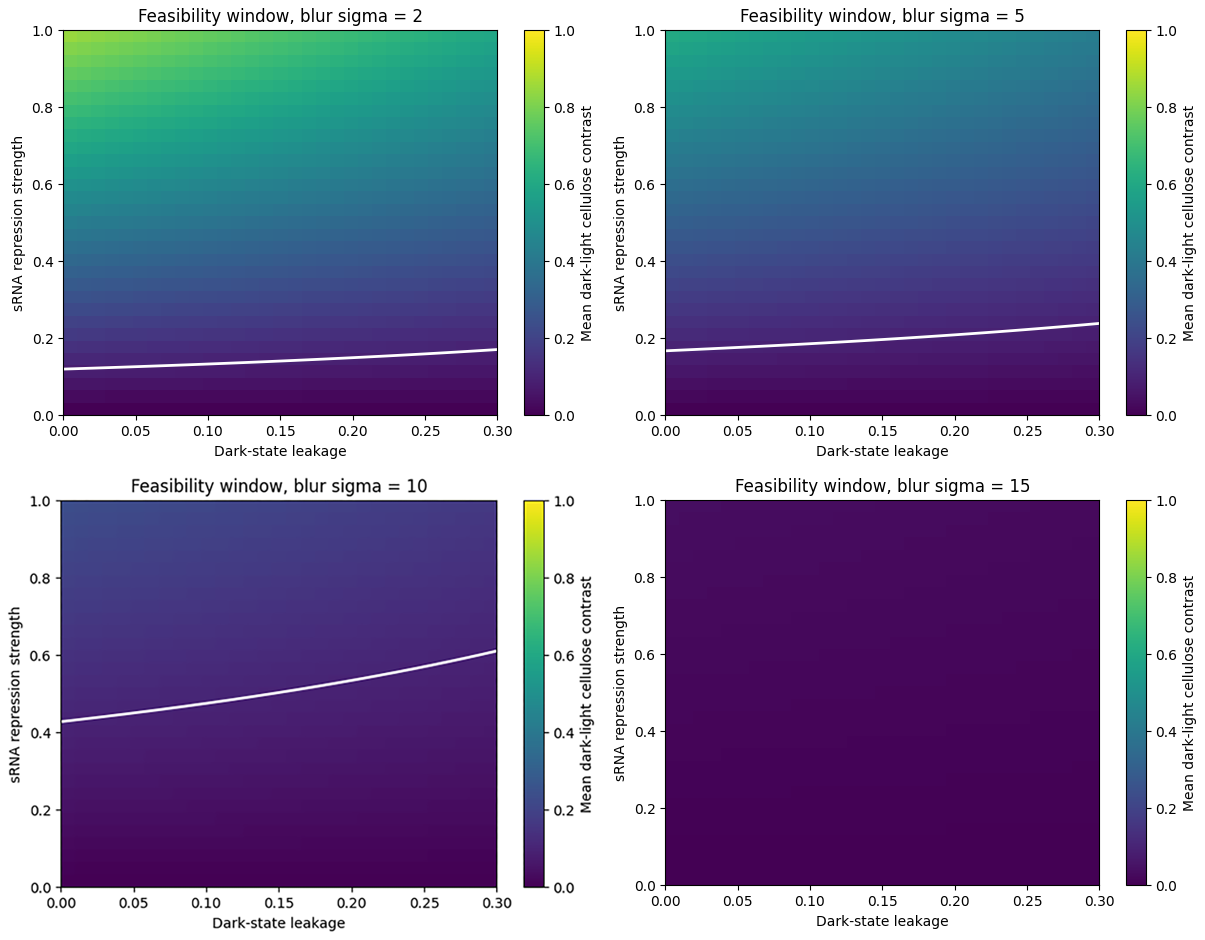 Effect of spatial blur on the feasibility window.
Effect of spatial blur on the feasibility window.
Increasing blur reduces the predicted feasibility window for visible cellulose patterning. This suggests that spatial resolution will depend not only on genetic circuit performance, but also on optical scattering, growth dynamics, and the physical behaviour of the cellulose pellicle. But it is worth noting that the leakage value is normalized between 0 and 1, not directly a biological percentage. As a design assumption, the model suggests that leakage should remain low, ideally below ~10–20% of ON-state response, while sRNA repression must be strong enough to overcome leakage. The exact repression threshold cannot be assigned biologically until the system is measured experimentally.”
 Stripe-width sensitivity test.
Stripe-width sensitivity test.
The same biological parameters were applied to different stripe widths. Narrower features are more easily lost under blur, while wider features remain more visible. This suggests that the final system would have a minimum pattern resolution below which material contrast may disappear.
The simulation suggests that visible cellulose patterning would require sufficiently low dark-state leakage and sufficiently strong sRNA-mediated repression. As spatial blur increases, the feasible region becomes smaller. This means that even if the genetic circuit functions, the material-level output could still fail if the biological or optical system blurs the pattern too strongly.
This simulation does not experimentally prove that optogenetic control and cellulose repression will work together in Komagataeibacter rhaeticus. Instead, it defines the parameter space that future wet-lab experiments would need to achieve.
Delay Model
The previous feasibility model assumed that the material response follows the projected light pattern immediately. However, the proposed biological system is not instantaneous. Light first activates the Opto-T7 system, which then drives sRNA production. The sRNA must accumulate, repress UGPase expression, and only then can cellulose production be affected.
To explore this, I created a simple delay model. A stripe pattern was shifted over time, and the cellulose output was calculated using either the current light pattern or a delayed version of the light pattern. The final material output was modelled as the average cellulose production over the simulated growth period.
The simulation shows that biological delay can reduce the fidelity of dynamic patterning. If the projected pattern changes faster than the cellular response time, the final cellulose output may become blurred or averaged. This suggests that future experimental patterning would likely require slow-changing or static light masks, at least in the first validation experiments.
This model does not provide a measured biological delay for the proposed system, but it highlights an important design constraint: the timing of light exposure must be matched to the response time of optogenetic activation, sRNA-mediated repression, and cellulose biosynthesis.
 Delay model
Delay model
When the biological response is assumed to be immediate, the final cellulose output follows the projected light pattern closely.
Figure. Delayed response model.
When a biological delay is introduced, the final cellulose pattern becomes an averaged record of earlier light exposure. This can reduce pattern sharpness and shift the material response away from the current projected pattern.
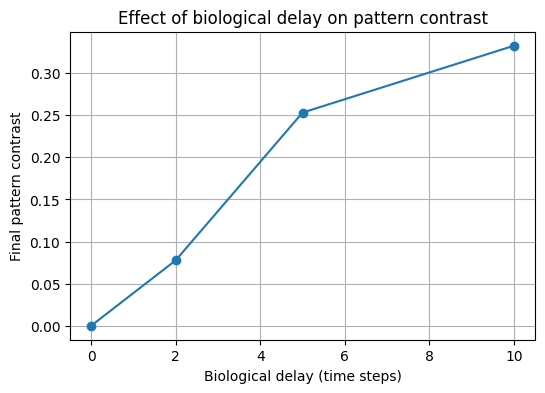 Effect of biological delay on predicted pattern contrast
Effect of biological delay on predicted pattern contrast
The model suggests that longer biological response delays can reduce the contrast of the final cellulose pattern, especially when the projected light pattern changes over time.
Literature on static Komagataeibacter BC growth often describes pellicle formation over days (5–15 days depending on strain and medium): for example, the top-fed BC reactor paper [8] delivered medium daily over 28 days, using 6 mL/day to cover the pellicle with about 1 mm of medium. Therefore it should be realistic to translate the simulation steps into 6 hours intervalls:
- 2 steps = 12 h
- 5 steps = 30 h
- 10 steps = 60 h / 2.5 days
A 10-step delay would then represent a slow material response of roughly 2–3 days, which matches the timescale where a pellicle becomes visibly established. The projection pattern should be held long enough for new cellulose to accumulate. As a first assumption, I would test 12- or 24-hour exposures, and treat multi-day exposures as the likely route to stronger material contrast.
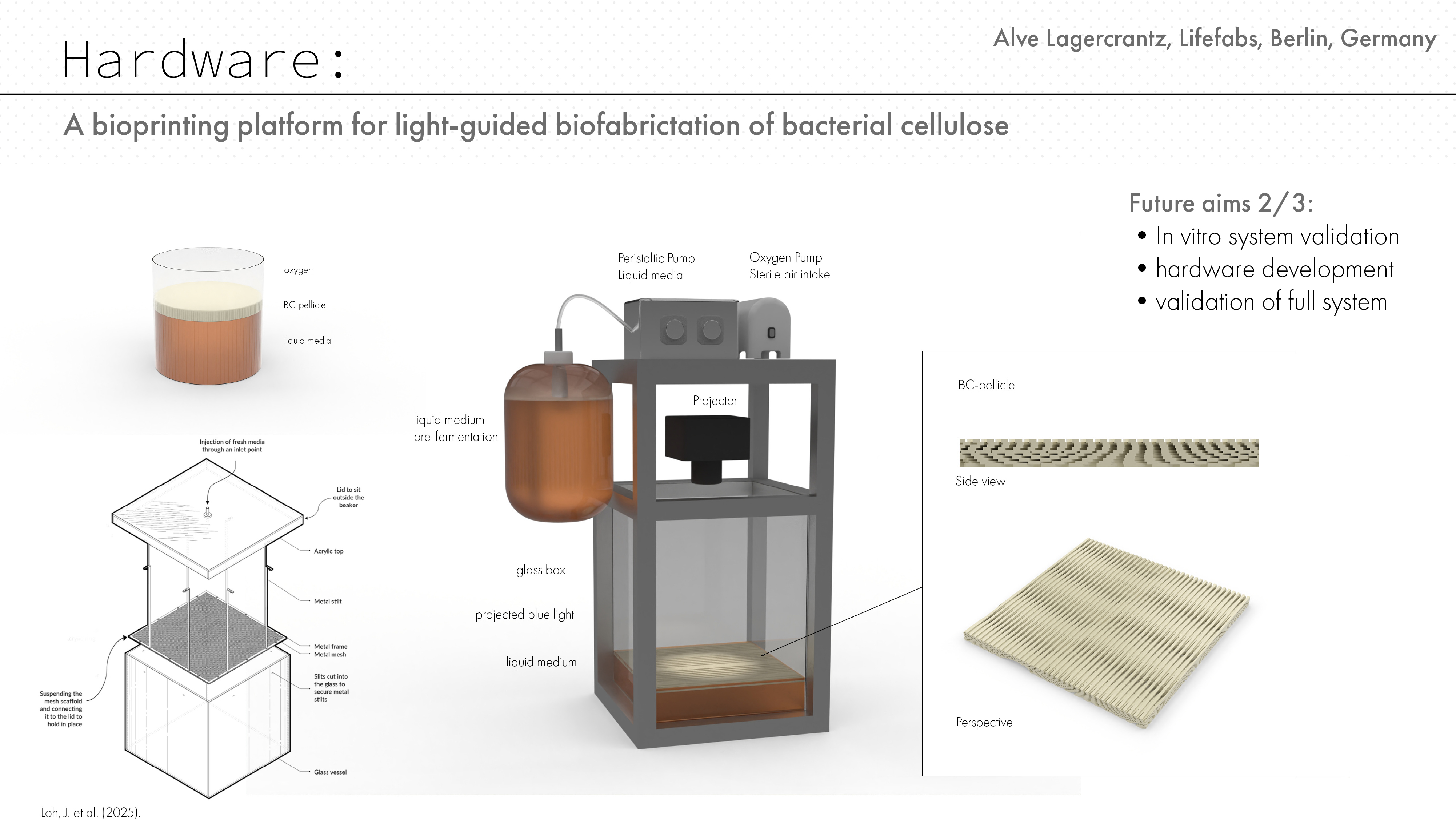
Hardware concept and visualisation
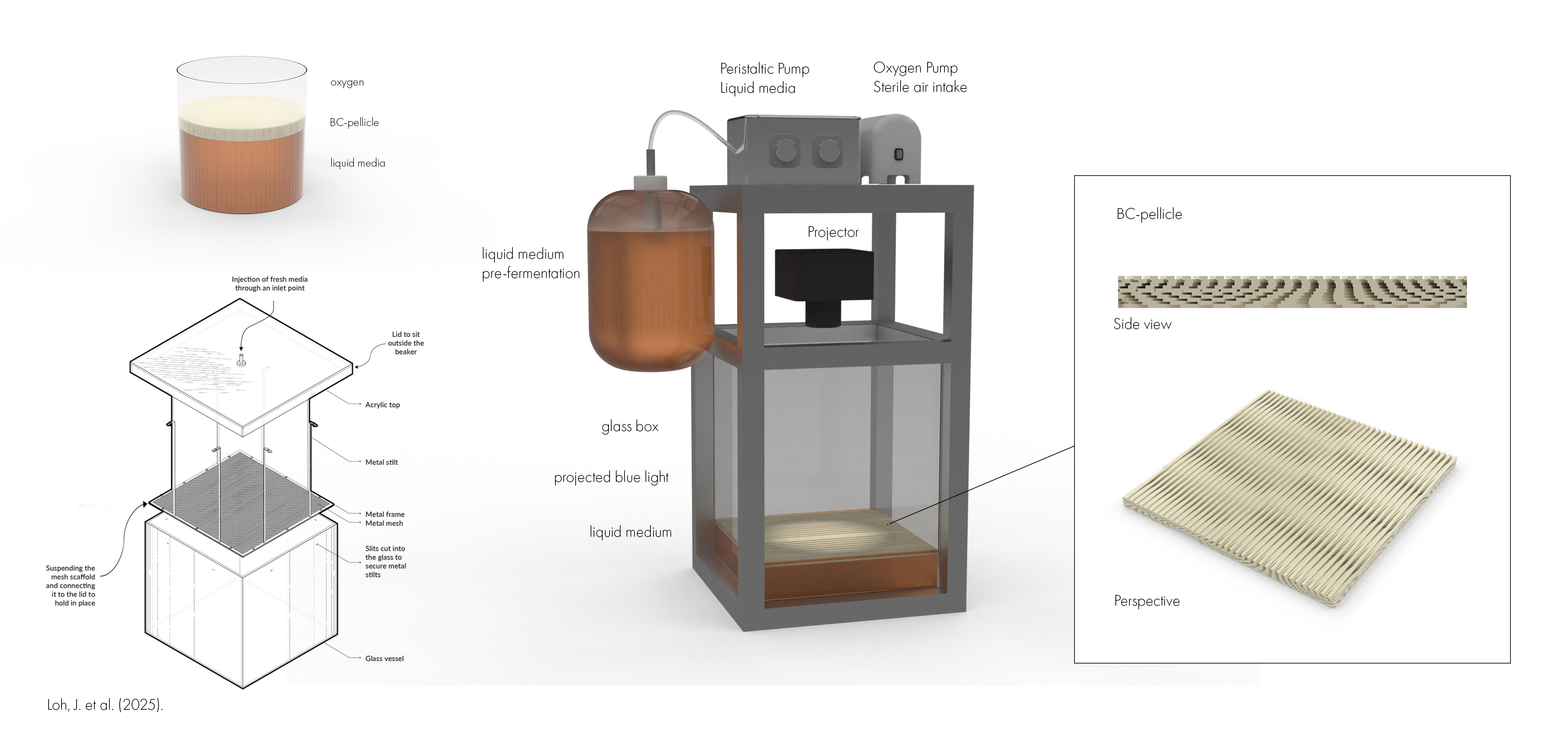 Hardware simulation
Hardware simulation
To explore how the proposed light-guided cellulose patterning system could become a physical platform, I developed a speculative hardware setup in Rhino. The render shows a small bioprinting chamber where Komagataeibacter grows bacterial cellulose at the air–liquid interface while a projected blue-light pattern controls local gene expression. The system combines a glass growth box, liquid medium reservoir, peristaltic medium delivery, sterile air/oxygen supply, and a projector positioned to expose the active growth surface. The design takes inspiration from the HBBE mesh-dispenser reactor described by Loh et al. (2025). Their system addresses a key limitation in static bacterial cellulose cultivation: as the pellicle becomes thicker, oxygen and nutrient access to the viable cells can become limited. By continuously or periodically supplying a thin layer of fresh medium to the pellicle surface, the reactor keeps the active growth zone better supported.
For my project, this design logic is especially useful because it could solve two problems at once. First, it helps maintain nutrient and oxygen access during long cellulose growth. Second, by encouraging growth at the upper surface, it reduces the need for projected light to pass through an increasingly thick cellulose sheet. This could make spatial light patterning more reliable over multi-day growth periods. The render is not a finished bioreactor, but a visual prototype for a future experimental setup. It connects the genetic circuit and simulation work to a possible material fabrication workflow, where bacterial cellulose could be grown with programmed spatial structure rather than produced as a uniform sheet.
9) Governance: safety, responsibility, and future use
This project is currently an in silico final project. No engineered Komagataeibacter rhaeticus strain was built, transformed, or grown as part of this work. However, the proposed future development would involve genetically engineered cellulose-producing bacteria, light-controlled gene expression, and possible material applications in textiles, fashion, and design. Because of this, the project raises governance questions around biosafety, environmental release, user safety, responsible communication, and open dissemination.
1. Governance and policy goals
The main governance goals for this project are:
A. Biosecurity
- Reduce the risk of inappropriate use or uncontrolled sharing of engineered organisms.
- Maintain traceability of genetic constructs, strains, and protocols.
- Share design knowledge responsibly without encouraging unsupervised release or unsafe replication.
B. Lab safety
- Ensure that future work with engineered K. rhaeticus happens only in appropriate lab settings.
- Use clear SOPs for strain handling, labeling, contamination response, and waste treatment.
- Avoid exposing users to live engineered organisms or unvalidated materials.
C. Environmental protection
- Prevent release of genetically modified organisms into the environment.
- Sterilize cultures, pellicles, and liquid waste before disposal.
- Evaluate whether final BC materials contain viable cells or residual genetic material before they leave the lab.
D. User and consumer protection
- Treat future textile or bag applications as skin-contact material applications.
- Test for sterility, leaching, irritation risk, wash stability, and durability before any public or wearable use.
- Clearly label experimental materials and avoid presenting speculative prototypes as consumer-ready products.
E. Feasibility, openness, and innovation
- Keep simulation notebooks, design logic, and hardware concepts open where possible.
- Avoid unnecessary restrictions that would prevent educational or artistic research.
- Share living strains and wet-lab protocols only through trained labs with appropriate biosafety infrastructure.
2. Governance actions
Option 1 — Contained research and strain-handling standard
Purpose:
Prevent accidental release or unsafe handling of engineered K. rhaeticus.
Design:
Future wet-lab work should use a containment-first approach. Engineered strains should be clearly labeled, stored, and documented. All cultures, pellicles, and liquid waste should be sterilized before disposal. No live engineered material should leave the lab. A simple strain registry should record the construct, host strain, plasmid/backbone, date, responsible person, and disposal status.
Assumptions:
The organism is not intended to be pathogenic, but it is genetically modified and should therefore be treated as a contained lab organism.
Risks:
Too much administrative complexity could make the project difficult for small labs or community labs. However, too little structure could lead to unsafe replication or accidental release.
Option 2 — Material safety and claim-labeling framework
Purpose:
Make sure that future BC materials are safe and honestly communicated before being used in textile, fashion, or bag applications.
Design:
Before any material is used outside the lab, it should be confirmed as non-living or appropriately inactivated. Future testing should include basic sterility checks, leaching tests, wash stability, skin-contact considerations, and mechanical/material performance. Documentation should clearly distinguish between:
- literature-supported claims
- in silico predictions
- experimental results
- speculative future applications
Assumptions:
Bacterial cellulose itself is often discussed as biocompatible, but engineered production systems, residual cells, media components, DNA, or processing chemicals could introduce additional risks.
Risks:
Testing requirements may slow down early-stage design experimentation. However, this is necessary if the material is intended for future skin-contact or consumer-facing applications.
Option 3 — Open-source but staged dissemination
Purpose:
Support open learning and reproducibility while avoiding uncontrolled distribution of engineered strains.
Design:
The project can openly share non-living and educational components, including:
- simulation notebooks
- design diagrams
- hardware concepts
- light-mask strategies
- governance documents
- general circuit logic
However, future wet-lab protocols, plasmids, and engineered strains should be shared more carefully, preferably through trained labs, institutional collaborations, or supervised workshops with biosafety infrastructure.
Assumptions:
Open-source biology can support education and innovation, but living engineered systems require more responsibility than digital files or passive materials.
Risks:
If too much is restricted, the project loses its educational and community-lab value. If too much is shared without context, people may copy the work without containment, testing, or waste procedures.
3. Specific risks for this project
| Risk | Why it matters | Governance response |
|---|---|---|
| Accidental release of engineered K. rhaeticus | The future organism would be genetically modified | Containment, labeling, sterilization, waste SOPs |
| Live cells remaining in final BC | Textile or bag materials may leave the lab | Sterility testing and inactivation before use |
| Overclaiming sustainability | BC is not automatically sustainable | Require transparent accounting of feedstock, energy, water, and processing |
| Overclaiming functionality | The system is not yet experimentally validated | Clearly separate simulated, literature-supported, and hypothetical claims |
| Unsafe community replication | Protocols could be copied without training | Staged dissemination and supervised sharing |
| Blue-light/projector hardware risks | Strong light, humidity, and electronics may create practical hazards | Enclosed hardware, eye protection, spill separation, electrical safety |
4. Score
Scoring system:
- 1 = strong / most favorable
- 2 = moderate
- 3 = weak / least favorable / highest burden
| Does the option: | Option 1 | Option 2 | Option 3 |
|---|---|---|---|
| Enhance Biosecurity | |||
| • By preventing incidents | 1 | 2 | 2 |
| • By helping respond | 1 | 2 | 2 |
| Foster Lab Safety | |||
| • By preventing incidents | 1 | 2 | 3 |
| • By helping respond | 1 | 2 | 3 |
| Protect the environment | |||
| • By preventing incidents | 1 | 2 | 3 |
| • By helping respond | 1 | 2 | 3 |
| Other considerations | |||
| • Minimizing costs and burdens to stakeholders | 2 | 3 | 1 |
| • Feasibility in community labs? | 3 | 2 | 1 |
| • Not impede research | 2 | 2 | 1 |
| • Promote constructive applications | 2 | 1 | 1 |
| Total | 15 | 20 | 20 |
Interpretation
Option 1 — Contained research and strain-handling standard
This option scores best overall because it most directly improves biosecurity, lab safety, and environmental protection. It is the strongest option for reducing risks associated with future work using engineered K. rhaeticus. Its main weakness is that it creates more burden and is less accessible for community-lab use.
Option 2 — Material safety and claim-labeling framework
This option is especially strong for responsible communication and future textile/material applications. It helps ensure that future bacterial cellulose materials are not overclaimed and are only presented as wearable or consumer-safe if tested appropriately. However, it adds significant testing and documentation burden.
Option 3 — Open-source but staged dissemination
This option performs best in terms of accessibility, research openness, and feasibility in community or educational settings. It supports constructive applications by keeping simulations, diagrams, and non-living design elements open. However, on its own it is weaker for direct biosafety and environmental protection.
4. Prioritization and recommendation
For this project, I would prioritize Option 1 and Option 2 as the baseline governance package.
Option 1 is necessary because future development would involve genetically engineered K. rhaeticus. The most important rule is that engineered strains and live cultures remain contained, traceable, and properly sterilized.
Option 2 is necessary because the long-term application is material production for textiles, fashion, and bags. Even if the biological system works, the material should not be treated as wearable or consumer-safe until it has been tested for viability, stability, leaching, and skin-contact safety.
Option 3 should be used as the dissemination strategy. The project should remain open and educational at the level of design logic, simulation, and hardware concepts, but living strains and wet-lab protocols should only be shared through trained labs or supervised collaborations.
5. Governance conclusion
The main governance challenge of this project is that it sits between biological research, material design, and future consumer-facing fabrication. The project is not currently a product and should not be presented as one. At this stage, the responsible claim is that the project proposes and simulates a possible route toward light-controlled bacterial cellulose structuring.
Future wet-lab development should therefore proceed step by step: first validating optogenetic reporter control, then testing sRNA-mediated UGPase repression, then measuring cellulose output, and only later exploring material applications. This staged approach allows the project to remain open, creative, and design-oriented while still respecting biosafety, environmental protection, and user safety.
10) Discussion and Future Work
- What is supported and what remains hypothetical
- Future wet-lab decision tree
- Main findings
- Drying/resolution note
What Is Supported and What Remains Hypothetical
| Claim | Status |
|---|---|
| K. rhaeticus can produce bacterial cellulose | Established |
| K. rhaeticus can be genetically engineered | Supported by literature |
| Opto-T7 can create light-controlled gene expression | Supported by literature |
| UGPase-targeting sRNA can reduce cellulose production | Supported by literature |
| PT7 can drive the UGPase-targeting sRNA cassette in this architecture | Proposed, not tested |
| Light-induced sRNA expression can reduce UGPase locally | Proposed, not tested |
| Local UGPase repression can create visible cellulose thickness/density patterns | Proposed, not tested |
| Layered moiré-like BC materials can be grown using this method | Visionary aim |
Future Wet-Lab Decision Tree (Aim 2)
Test Opto-T7 with a fluorescent reporter.
- If light/dark contrast is low: tune arabinose, promoter strength, or plasmid copy number.
- If contrast is good: continue.
Replace reporter with PT7-sRNA and measure sRNA expression.
- If sRNA is not light-induced: redesign the output cassette.
- If sRNA is light-induced: continue.
Measure UGPase repression.
- If UGPase is not reduced: redesign the sRNA or Hfq expression strategy.
- If UGPase is reduced: continue.
Measure cellulose output.
- If cellulose does not change: UGPase repression is insufficient or cellulose regulation is buffered.
- If cellulose changes globally: leakage is too high.
- If cellulose changes locally: proceed to spatial patterning.
Test simple light masks.
- Start with large stripes or blocks.
- Only later test fine patterns or moiré layers.
Main Findings from the In Silico Feasibility Study
The simulations suggest four main design constraints for future optogenetic cellulose patterning:
Model-based feasibility window
- Resolution: 0.8 mm is the predicted lower visible limit in the model; first wet-lab tests should start with safer ~2 mm stripe features.
- Leakage: dark-state activity should remain low, ideally below 10–20% of ON-state response, to preserve contrast between exposed and unexposed regions.
- Blur: effective biological blur must stay smaller than the feature width; in the model, 0.2 mm features disappear, 0.8 mm features survive, and 2 mm features remain strong.
- Repression: anti-UGPase sRNA must create sufficient knockdown to overcome leakage and produce a measurable reduction in cellulose biosynthesis.
- Delay / projection rate: cellulose is a slow material output, not an instant reporter. First tests should use sustained projections, for example 12–24 h exposures, with stronger contrast likely requiring multi-day patterning.
I use 0.8 mm as a reference from the optogenetic paper, but I do not claim this will be the first achievable cellulose resolution. Because cellulose is a growing material output, I expect the first realistic patterns to be larger stripes or gradients. The first experimental validation should use simple, large-scale patterns such as stripes or blocks, not high-resolution images or moiré patterns. It is also worth noting that in this project we have only been discussing resolution in wet state but BC lose 80-90 percent of its volume during drying which will completly change the resolution of the final product.
What Was Achieved in This Project
This project did not build the biological system experimentally. It produced a complete in silico design package: proposed circuit architecture, synthesis-ready DNA fragments, Twist feasibility check, Google Colab simulations, a Rhino hardware concept, and a staged wet-lab validation plan.
The achieved outcomes are:
- Designed a proposed genetic architecture linking blue-light Opto-T7 control to sRNA-mediated UGPase repression.
- Designed four synthesis-ready DNA fragments: OptoT7 half 1, OptoT7 half 2, PT7-sRNA, and Hfq helper cassette.
- Checked commercial synthesis feasibility using Twist Bioscience.
- Built spatial simulations linking light input, dark-state leakage, repression strength, blur, and predicted cellulose output.
- Estimated how feature size affects pattern retention.
- Simulated how biological delay affects dynamic patterning.
- Defined the key wet-lab validation steps needed to test the system experimentally.
Checked techniques:
- Bioethical considerations
- DNA construct design
- Databases: Addgene, GenBank / NCBI, Benchling
- Creating Twist order
- Other cloning methods: Golden Gate assembly / KTK, future work
- Chassis selection: K. rhaeticus
- Bacterial culturing: future Aim 2
- Quality control / analysis: fluorescence, OD600, cellulose output
- Computational analysis: Python / Google Colab
- Hardware design: Rhino / bioreactor concep
DNA construct design:
In this project, DNA construct design was used to convert the proposed biological logic into synthesis-ready modules. I designed separate OptoT7 half 1 and half 2 input cassettes, a PT7-controlled anti-UGPase sRNA output cassette, and an Hfq helper cassette. These constructs were checked for orientation, regulatory architecture, synthesis constraints, and Twist compatibility. This validates that the proposed system can be translated from a conceptual circuit into physical DNA fragments.
Computational analysis:
Computational analysis was used to test whether projected light patterns could theoretically remain visible after biological leakage, repression strength, blur, and delay. Python simulations converted light inputs into predicted cellulose-output maps and compared different stripe widths, blur levels, and leakage values. The results do not prove wet-lab function, but they define a design window for future experiments. This helped identify that first tests should use simple millimetre-scale stripes rather than fine images or moiré patterns.
11. References
[1] Arnardottir, T. H. et al. Living Manufacture: Principles for a microbial 3D printer. download
[2] Goosens, V. J. et al. (2021).
https://doi.org/10.1021/acssynbio.1c00358
[3] Teh, M. Y. et al. (2019).
https://doi.org/10.1021/acssynbio.8b00168
[4] Walker, K. T. et al. (2025).
https://www.nature.com/articles/s41587-024-02194-3
[5] Levskaya, A. et al. (2005).
https://doi.org/10.1038/nature04405
[6] Florea, M. et al. (2016).
https://doi.org/10.1073/pnas.1522985113
[7] Zhou, H. et al. (2026).
https://doi.org/10.1016/j.tibtech.2025.09.019
[8] Loh, J. et al. (2025).
https://link.springer.com/article/10.1007/s10570-024-06367-w
[9] Cielecka, I. et al. (2020).
https://www.mdpi.com/2076-3417/10/11/3850
Full Zotero reff list can be found here: