Week 11: Bioproduction & Cloud Labs
Week 11: Bulding Genomes

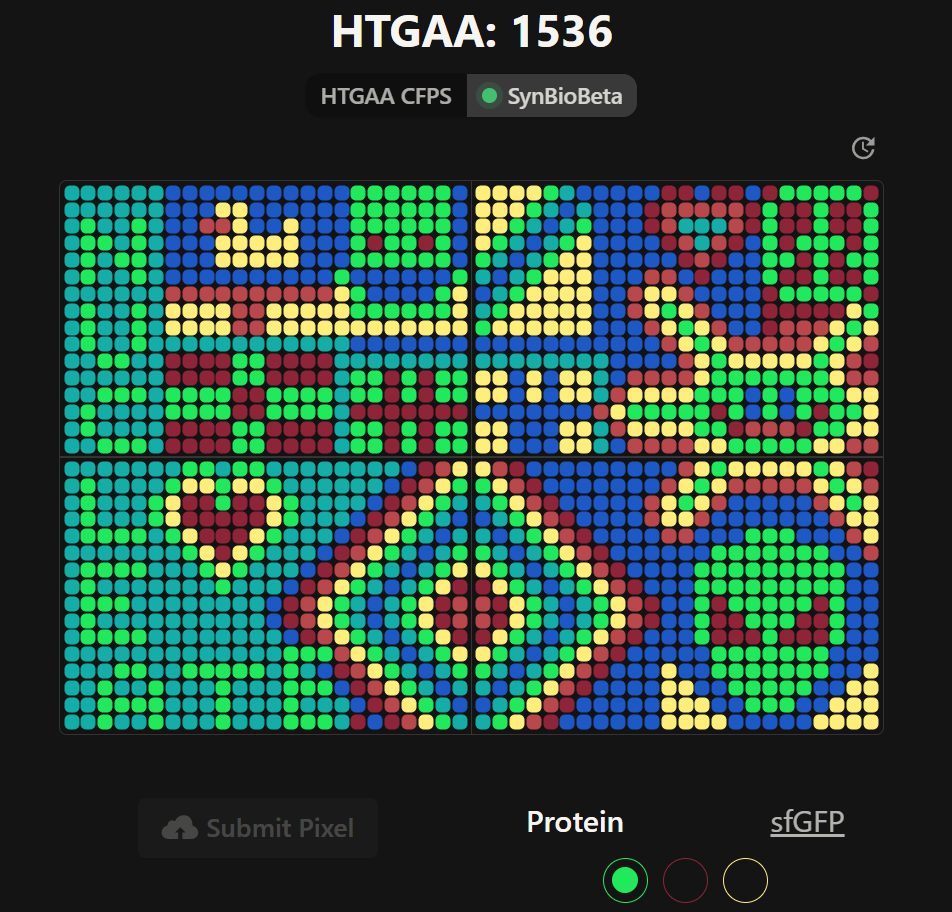
Part A: The 1,536 Pixel Artwork Canvas | Collective Artwork
Pixel Artwork Contributation:
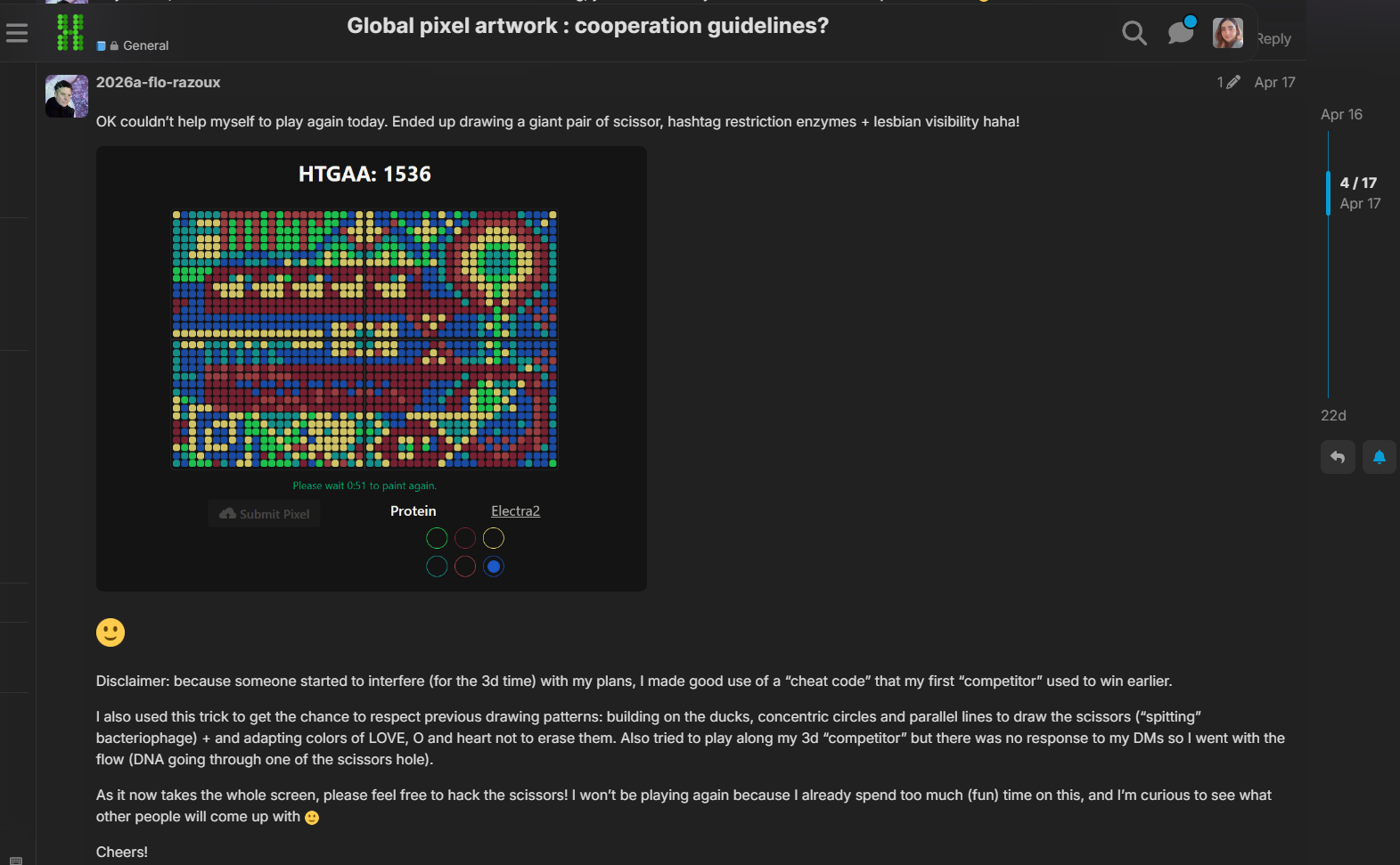
I was excited to participate in the HTGAA 2026 collaborative pixel artwork experiment. My main contribution was helping create part of the “2026” design on the left corner of the canvas, as well as contributing to the yellow and cyan giraffe section, the rainbow mandala near the center-bottom region, and a small Ecuadorian flag on the left side of the artwork showed on the Figures 1 and Figure 7. The collaborative work was on: https://rcdonovan.com

Figure 1. My contributions to the HTGAA 2026 collaborative pixel artwork project. Made on April 19th 2026
| Before | After |
|---|---|
| 
|
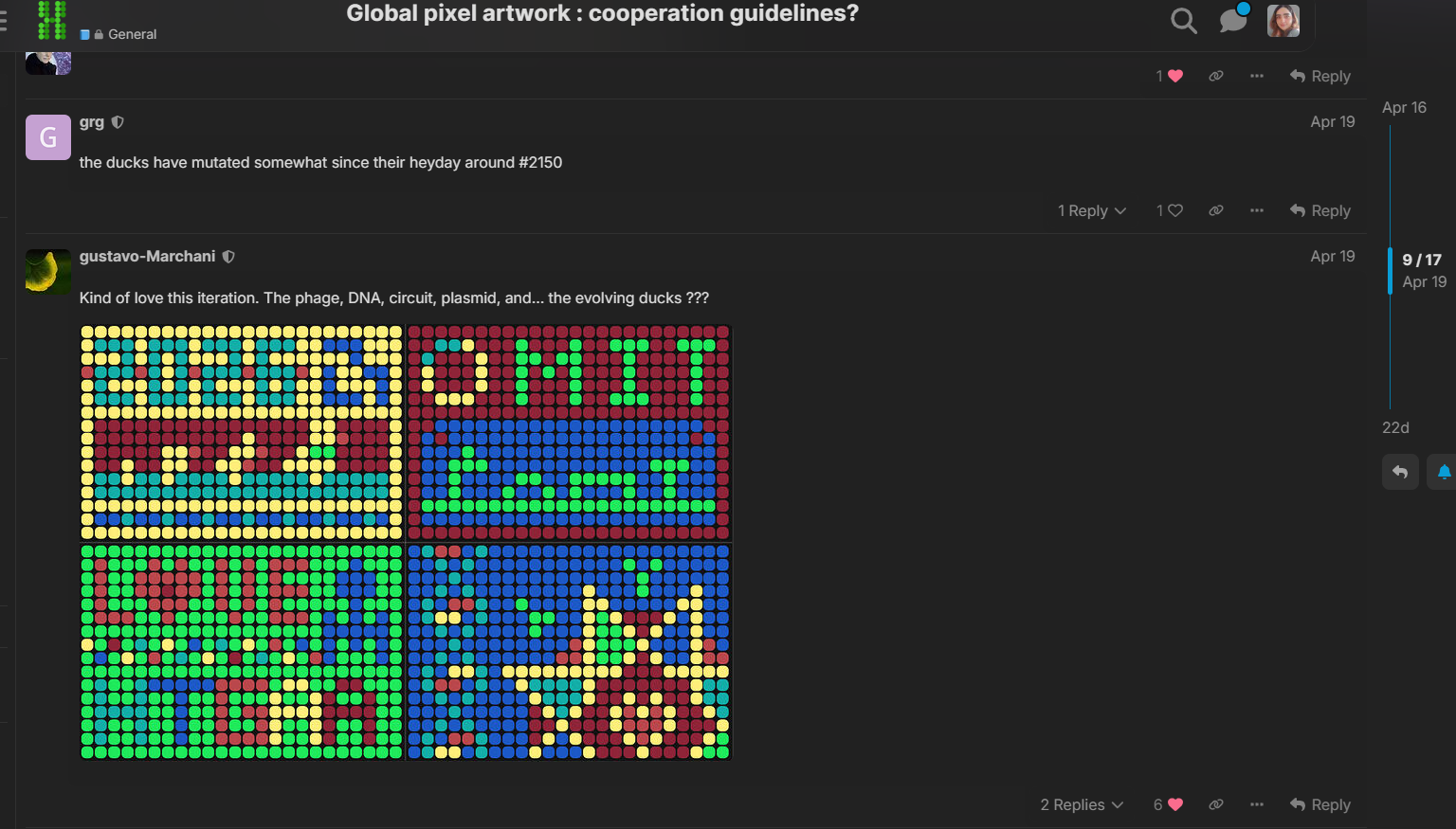
Figure 2. Comparison between before and after by other global students
Figure 3. Video of the Timeline made by Ronan!
Link of the Art Forum: HTGAA Forum
One of the things I enjoyed the most about this project was seeing how students from different countries and backgrounds collaborated in real time to build a single large-scale bioart piece together. I especially liked browsing the HTGAA forum and observing how creative and diverse the ideas from other global participants were (Figure 2). Additionally, the timeline visualization created by Ronan using the pixel history tool made the collaborative process feel even more dynamic and memorable (Figure 3).
I also appreciated how this project combined synthetic biology, digital art, and community participation into a shared scientific-artistic experience. It made the course feel more connected globally, even for remote participants and committed listeners! Finally, for the collab of HTGAA 2026 and SynBioBeta, the final image of the global collab image (Figures 6 & 7).
Transition to the reagent collaboration section:
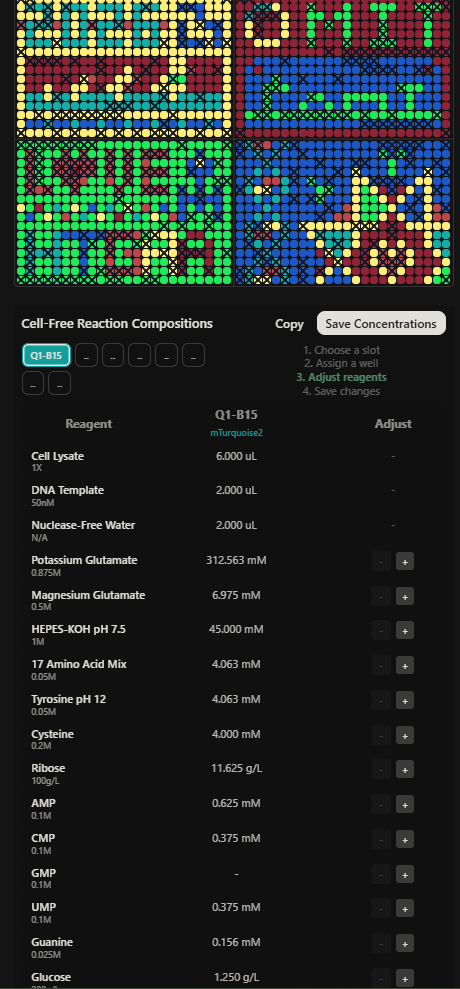
As part of the second collaborative phase of the experiment, I also contributed to preliminary reagent composition planning for the cell-free fluorescent artwork reactions. My contribution focused on testing small variations in ribose and glucose supplementation across multiple wells in order to explore how sustained energy metabolism could affect long-term fluorescence during extended incubation (Figures 4 & 5).
Gallery:
| Figure 4 | Figure 5 |
|---|---|
| 
|
Figure 4. Initial reagent adjustment planning for cell-free fluorescence optimization; Figure 5. Table of contribution ana-gomez
Output code:
▶ Reagent Supplement JSON (Click to Open)
| 
|
|---|
Figure 6. Final collaborative HTGAA 2026 fluorescent artwork. HTGAA 2026 (May 20, 2026). Most recent Art pixel collaboration screenshot
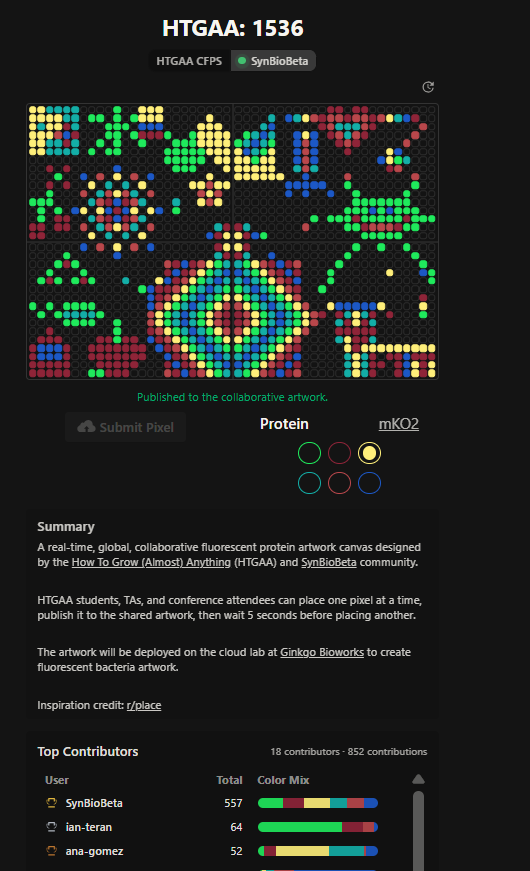
Figure 7. HTGAA 2026 collaborative artwork before the SynBioBeta integration (May 05, 2026). I contributed to the giraffe section on the left corner with the yellow and cyan background, the rainbow mandala near the bottom center, and the small Ecuadorian flag on the left side of the canvas.
Future recommendations: I think it could be improved by providing a larger editable canvas area or allowing teams to reserve small collaborative regions in advance. This could help participants coordinate more complex designs while still maintaining the spontaneous and creative nature of the project. Additionally, one aspect that noticeably improved during the experiment was the pixel placement cooldown time, which was reduced from 30 seconds at the beginning to only 5 seconds later on. This significantly improved the collaborative experience and allowed participants to contribute more efficiently to the evolving artwork.

Part B: Cell-Free Protein Synthesis | Cell-Free Reagents
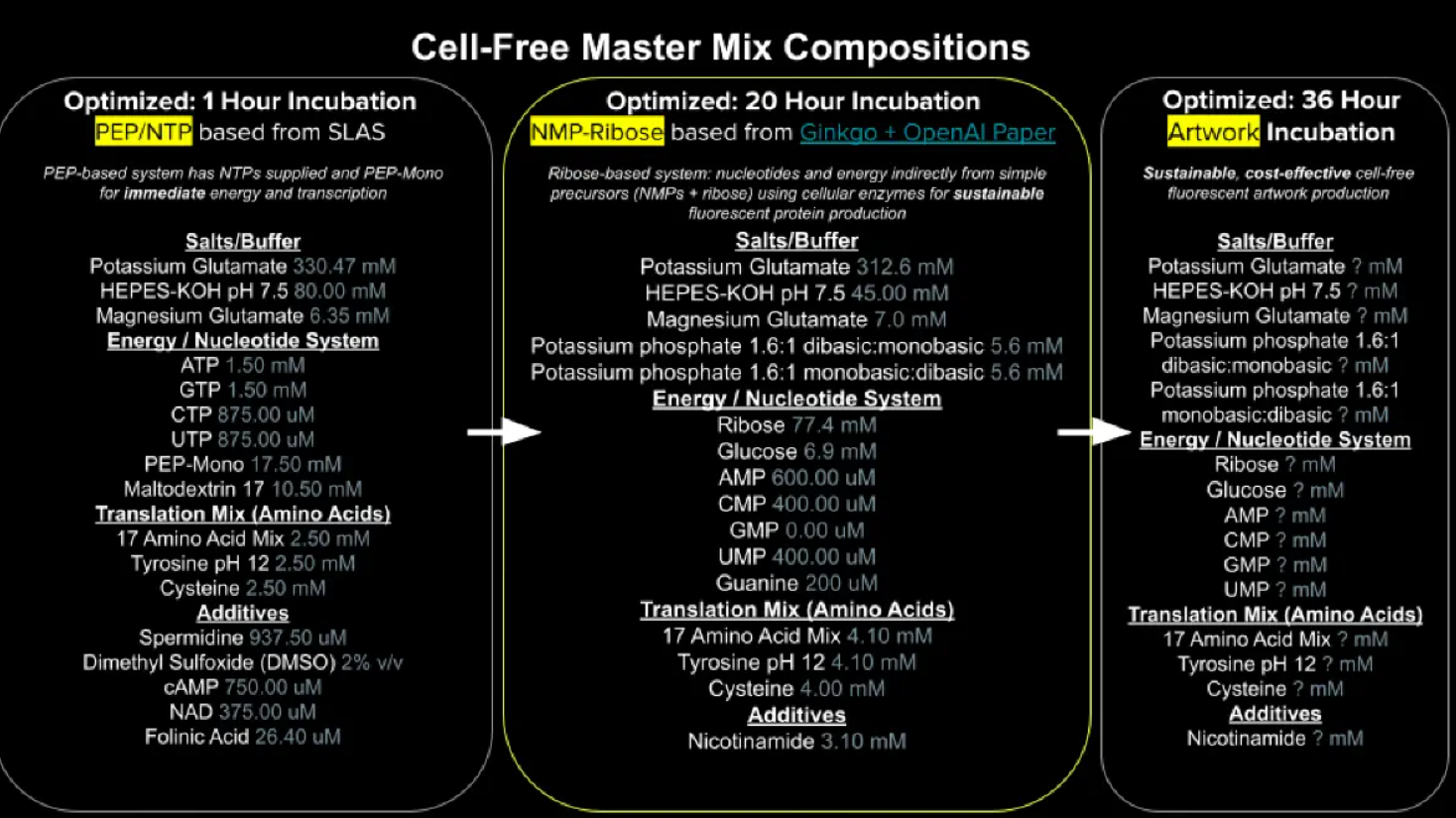
Figure 8. Cell Free Master Mix-Composition | MIT-HTGAA 2026 | Week 11: HTGAA 2026 WEEK 11
The optimized 20-hour incubation formulation is a ribose-based cell-free TXTL system designed to sustain long-duration fluorescent protein expression through indirect nucleotide and energy regeneration pathways. The reaction composition can be divided into the following categories: E. coli Lysate, Salts/Buffer, Energy/Nucleotide System, Translation Mix (Amino Acids), Additives, and Backfill.
1. E. coli Lysate
BL21 (DE3) Star Lysate (includes T7 RNA Polymerase)
- This lysate contains the endogenous E. coli transcription-translation machinery required for cell-free protein synthesis, including ribosomes, enzymes, tRNAs, metabolic cofactors, and ATP regeneration pathways.
- The incorporated T7 RNA polymerase enables strong transcription from T7 promoters commonly used in synthetic biology, although its high transcriptional speed may affect transcription-translation coupling under some reaction conditions.
(Silverman et al., 2020; Zhang et al., 2018)
2. Salt / Buffers
| Reactive | Description | References |
|---|---|---|
| Potassium Glutamate | Maintains ionic strength and mimics the intracellular E. coli environment, supporting ribosome stability and protein synthesis. It was consistently retained across optimized RFopt TXTL formulations. | (Deredge et al., 2010; Olsen et al., 2026) |
| HEPES-KOH pH 7.5 | Stabilizes pH during long incubations, helping maintain transcription and translation efficiency. HEPES supplementation also improved long-duration RFopt performance. | (Panah et al., 2015; Olsen et al., 2026) |
| Magnesium Glutamate | Essential for ribosome assembly, RNA stability, and ATP-dependent enzymatic reactions during transcription and translation. It remained a key component in optimized TXTL systems. | (Caschera, 2017; Sun et al., 2023; Olsen et al., 2026) |
| Potassium phosphate monobasic/dibasic | Supports phosphate balance, buffering capacity, and metabolic stability during TXTL reactions. Phosphate levels also influenced ATP balance in RFopt formulations. | (Anderson et al., 2015; Yadav et al., 2025; Olsen et al., 2026) |
3. Energy / Nucleotide System
| Reactive | Description | References |
|---|---|---|
| Ribose | Supports nucleotide regeneration and sustained transcriptional activity during long-duration TXTL reactions. Ribose supplementation was also associated with improved yields in optimized RFopt formulations. | (Loan et al., 2018; Olsen et al., 2026) |
| Glucose | Functions as a low-cost non-phosphorylated energy substrate that supports ATP regeneration and prolonged TXTL activity in E. coli lysate systems. | (Anderson et al., 2015; Sato et al., 2024; Olsen et al., 2026) |
| AMP / CMP / UMP | These nucleotide monophosphates can be recycled into higher-energy nucleotide forms required for RNA synthesis and metabolic maintenance during cell-free protein synthesis. | (Nakagawara et al., 2022) |
| GMP | Contributes to guanosine nucleotide pools required for transcription and RNA synthesis. Optimized RFopt systems demonstrated that GMP could be partially replaced using guanine and ribose supplementation. | (Ballut et al., 2023; Olsen et al., 2026) |
| Guanine | Guanine can be converted into GMP and eventually GTP through endogenous nucleotide salvage pathways present in the lysate. Guanine and ribose supplementation successfully replaced GMP in optimized RFopt formulations without major yield reduction. | (Olsen et al., 2026; Wang et al., 2022; Iglesias-Gato et al., 2010) |
4. Translation Mix
| Reactive | Description | References |
|---|---|---|
| 17 Amino Acid Mix | Provides most amino acids required for translation, protein synthesis, and fluorescent protein production during TXTL reactions. Amino acid composition and concentration are important for maintaining efficient cell-free expression systems. | (Caschera & Noireaux, 2015; Worst et al., 2016) |
| Tyrosine | Tyrosine is supplemented separately due to its lower solubility and potential depletion during long-duration TXTL reactions. Separate preparation can improve reagent stability and protein expression efficiency. | (Caschera & Noireaux, 2015; Thornton et al., 2025) |
| Cysteine | Cysteine is added separately because of its oxidation sensitivity and its role in protein folding and disulfide bond formation. Redox optimization is particularly important for cysteine-rich proteins in cell-free systems. | (Ruiz et al., 2022) (Siddiquee & Kwan, 2021) |
5. Additives
Nicotinamide:
- Supports redox balance and NAD-related cofactor regeneration pathways involved in metabolic activity during TXTL reactions and cell-free protein synthesis.
(Alegre & Pastore, 2023; Yusri et al., 2025)
6. Backfill
Nuclease Free Water:
- Used to adjust reaction volumes while minimizing nucleic acid degradation during TXTL and cell-free expression reactions.
(Waller, 2024; Romantseva et al., 2022)
Useful Resources for Cell-Free Reaction Preparation:
Online buffer calculators can help estimate reagent compositions during cell-free master mix preparation and optimization workflows. One useful example is the AAT Bioquest HEPES buffer preparation calculator.
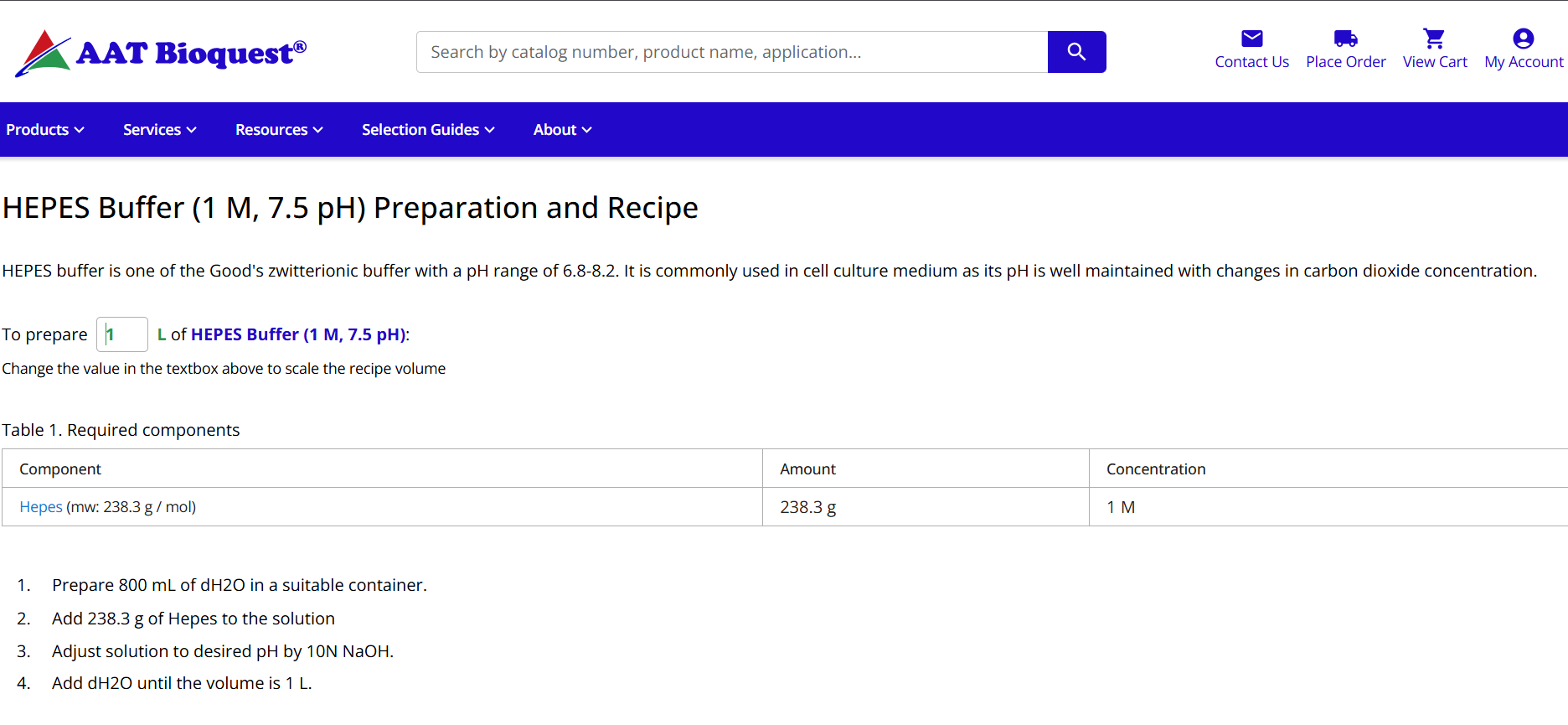
| https://www.aatbio.com/resources/buffer-preparations-and-recipes/hepes-buffer-ph-7-5 |
Figure 9. AAT Bioquest Online Calculator | HEPES Buffer
Difference Between Systems
1-hour optimized PEP-NTP master mix vs. 20-hour NMP-Ribose-Glucose master mix
- The 1-hour PEP-NTP system is optimized for rapid and high initial protein expression by directly supplying nucleotide triphosphates and phosphoenolpyruvate (PEP) as an immediate energy source. In contrast, the 20-hour NMP-ribose-glucose system relies on slower metabolic recycling pathways using nucleotide monophosphates, ribose, and glucose, creating a more sustainable and cost-effective platform for prolonged fluorescent protein expression and bioart applications.
- Recent studies further demonstrated that optimized RFopt formulations maintained ATP levels for longer periods, reduced phosphate accumulation, and supported a more balanced metabolic environment during extended TXTL reactions. (Olsen et al., 2026)
Bonus Question
How can transcription occur if GMP is not included, but Guanine is?
Transcription can still occur because enzymes present in the E. coli lysate can convert guanine into GMP and eventually GTP through endogenous nucleotide salvage pathways, enabling continued RNA synthesis even without externally supplied GMP. Supporting this concept, Olsen et al. (2026) demonstrated that a combination of guanine and ribose could successfully replace GMP in optimized RFopt formulations without significantly reducing protein yield.

Part C: Planning the Global Experiment | Cell-Free Master Mix Design
For the reagent optimization step, I introduced only minor adjustments to ribose and glucose concentrations while maintaining the remaining core TXTL components unchanged. These reagents were selected because they primarily contribute to energy regeneration and long-term metabolic support in the cell-free system, allowing subtle modulation of fluorescence performance without significantly disrupting transcriptional or translational stability.
In contrast, key components such as magnesium glutamate, potassium glutamate, HEPES buffer, amino acid mixtures, and the E. coli lysate was intentionally preserved at their optimized baseline concentrations due to their essential roles in maintaining ionic balance, pH stability, ribosomal activity, protein synthesis efficiency, and overall reaction robustness. Limiting modifications to energy-associated reagents reduced the risk of compromising fluorescence output or affecting reproducibility across the collaborative cloud-lab experiment.
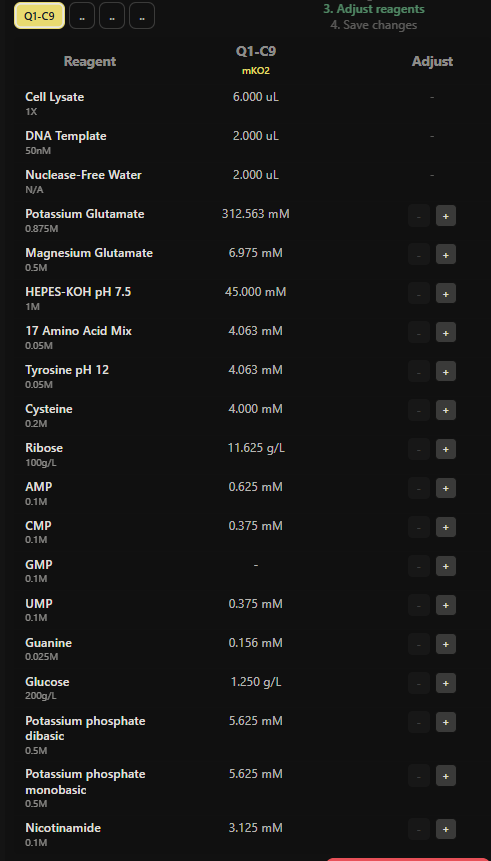
Figure 10. Master mix composition and reagent adjustment workflow for Q1-C9 expressing mKO2 during the HTGAA 2026 collaborative cell-free experiment.
Reagent Adjustment Strategy
The reagent supplementation strategy focused on testing small variations in ribose and glucose concentrations across selected wells while balancing total reaction volume with nuclease-free water. These modifications were designed to explore how sustained metabolic support and ATP regeneration could influence long-duration fluorescent protein expression during the 36-hour incubation period.
▶ Reagent Supplement JSON (Click to Open)
Fluorescent Proteins:
| Fluorescent Protein | Biophysical / Functional Property | References |
|---|---|---|
| sfGFP | sfGFP is highly robust because it folds efficiently and matures rapidly, making it reliable for cell-free fluorescent readouts. | (Pédelacq et al., 2006; Pharma, 2025; FPbase, 2022) |
| mRFP1 | mRFP1 is useful as a red fluorescent readout, but its brightness and maturation behavior can affect how quickly fluorescence becomes detectable. | (Balleza et al., 2017; FPbase, 2002) |
| mKO2 | mKO2 fluorescence can be influenced by oxygen availability and maturation kinetics, which may affect signal intensity during extended cell-free incubation. | (Kremers et al., 2009; FPbase, 2008) |
| mTurquoise2 | mTurquoise2 is a bright cyan fluorescent protein with high quantum yield, fast maturation, and low acid sensitivity, making it suitable for long-duration fluorescence measurements. | (Goedhart et al., 2012; FPbase, 2021) |
| mScarlet_I | mScarlet-I is a bright monomeric red fluorescent protein with relatively low pH sensitivity, which may help maintain fluorescence during long TXTL reactions. | (Botman et al., 2019; FPbase, 2016) |
| Electra2 | Electra2 is a blue fluorescent protein derived from Entacmaea quadricolor; as a blue FP, its readout may depend strongly on proper folding, maturation, and spectral separation from green/red fluorescent proteins. | (Papadaki et al., 2022; FPbase, 2022) |
Hypothesis
I hypothesize that supplementing reactions expressing mKO2 and mScarlet-I with moderate additional ribose and glucose could improve fluorescence after a 36-hour incubation by sustaining ATP regeneration and prolonged protein synthesis. Since fluorescent protein output depends not only on expression but also on maturation efficiency and reaction stability, maintaining metabolic activity may increase total fluorescent protein accumulation and final fluorescence intensity.
This hypothesis is inspired by RFopt-style reagent optimization approaches, where ribose-containing formulations and balanced metabolic activity supported improved long-duration TXTL performance.

Part D: Build-A-Cloud-Lab | (optional) Bonus Assignment

Figure 11. Automated cell-free fluorescence screening and routing system for TXTL optimization designed using the Ginkgo Nebula cloud-lab simulation tool. The conceptual layout includes sample intake, quality-control analysis, TXTL processing modules, fluorescence analysis, automated routing systems, and distributed storage infrastructure connected through modular transport components.
The cloud-lab layout was conceptually designed as an automated cell-free fluorescence screening and routing system for TXTL optimization. The modular workflow integrates sample intake, quality-control analysis using Lunatic instrumentation, TXTL processing stations, fluorescence analysis modules, automated routing pathways, and distributed storage infrastructure connected through transport components and switching systems.
The additional routing pathway was included to conceptually represent sample reanalysis or rerouting when quality-control parameters or fluorescence outputs fail to meet expected thresholds. This design explores how future cloud laboratories could integrate modular automation, distributed experimentation, and autonomous workflow decision-making into scalable synthetic biology research environments.
Additionally, the following table summarizes the conceptual function of each component used in the cloud-lab workflow design.
| Model | Type | Characteristics | References |
|---|---|---|---|
| Nebula | System | Modular robotic biotechnology workflow infrastructure with interconnected units for cloud-lab processing, routing, and distributed storage systems. | (Ginkgo Bioworks, 2026; Bioworks, 2026) |
| Lunatic | Instrument | Instrument used for DNA/RNA quantification, protein analysis, and absorbance-based quality control measurements. | (Wintermute, 2024; Bioworks, 2026) |
| RAC-E15 / E16 / E17 | Component | Reconfigurable robotic automation carts of different sizes used for modular experimental workflows and transport integration. | (Wintermute, 2024; Ginkgo Bioworks, 2026) |
| MM-Turn | Component | Modular connection unit used to redirect or rotate transport pathways between workflow modules. | (Wintermute, 2024; Bioworks, 2026) |
| MM-0.25m / MM-1m | Component | Modular transport/interconnect segments with lengths of 0.25 m and 1 m used to connect workflow stations. | (Wintermute, 2024; Bioworks, 2026) |
| MM-Switch | Component | Routing and switching module used to redirect samples between alternative workflow pathways. | (Wintermute, 2024; Bioworks, 2026) |
| Taxi (5-puck) | Component | Automated transport component designed for the movement of multiple samples, plates, or puck-based carriers between workflow stations. | (Wintermute, 2024; Bioworks, 2026) |
Some functional interpretations were conceptually inferred from the cloud-lab simulation environment used to generate Figure 11 and may not represent the complete real-world implementation of the Ginkgo RAC infrastructure.

Weekly Reflection
- This week helped me better understand how small reagent modifications can influence long-duration TXTL fluorescence performance.
- I really liked exploring the cloud-lab simulator because it made me think about how future synthetic biology workflows could be automated and modular.
- One thing that surprised me was how much reaction stability and metabolic balance matter for maintaining fluorescence over time.
- Designing the rerouting pathway in the cloud-lab workflow made me think more about quality control and autonomous experimental systems.
- I enjoyed how this week combined bioart, automation, and synthetic biology into a collaborative experiment.
Thanks for reading! You can find this info in my personal website in Notion → Link: Notion: Week 11-Bioproduction & Cloud Labs

References and Sources
References:
Part B:
Alegre, G. F. S., & Pastore, G. M. (2023). NAD+ Precursors Nicotinamide Mononucleotide (NMN) and Nicotinamide Riboside (NR): Potential Dietary Contribution to Health. Current nutrition reports, 12(3), 445–464. https://doi.org/10.1007/s13668-023-00475-y
Anderson, M. J., Stark, J. C., Hodgman, C. E., & Jewett, M. C. (2015). Energizing eukaryotic cell-free protein synthesis with glucose metabolism. FEBS letters, 589(15), 1723–1727. https://doi.org/10.1016/j.febslet.2015.05.045 | NCBI: https://pmc.ncbi.nlm.nih.gov/articles/PMC4651010/
Ballut, L., Violot, S., Kumar, S., Aghajari, N., & Balaram, H. (2023). GMP Synthetase: Allostery, Structure, and Function. Biomolecules, 13(9), 1379. https://doi.org/10.3390/biom13091379
Caschera, F. (2017). Bacterial cell-free expression technology to in vitro systems engineering and optimization. Synthetic and Systems Biotechnology, 2(2), 97–104. https://doi.org/10.1016/j.synbio.2017.07.004
Caschera, F., & Noireaux, V. (2015). Preparation of amino acid mixtures for Cell-Free Expression Systems. BioTechniques, 58(1), 40–43. https://doi.org/10.2144/000114249
Deredge, D. J., Baker, J. T., Datta, K., & Licata, V. J. (2010). The glutamate effect on DNA binding by pol I DNA polymerases: osmotic stress and the effective reversal of salt linkage. Journal of molecular biology, 401(2), 223–238. https://doi.org/10.1016/j.jmb.2010.06.009 [No Open Access]
Iglesias-Gato, D., Martín-Marcos, P., Santos, M. A., Hinnebusch, A. G., & Tamame, M. (2011). Guanine nucleotide pool imbalance impairs multiple steps of protein synthesis and disrupts GCN4 translational control in Saccharomyces cerevisiae. Genetics, 187(1), 105–122. https://doi.org/10.1534/genetics.110.122135
Loan, T. D., Easton, C. J., & Alissandratos, A. (2018). Recombinant cell-lysate-catalysed synthesis of uridine-5′-triphosphate from nucleobase and ribose, and without addition of ATP. New Biotechnology, 49, 104–111. https://doi.org/10.1016/j.nbt.2018.10.002 [No Open Access]
Nakagawara, K., Takeuchi, C., & Ishige, K. (2022). 5’-CMP and 5’-UMP promote myogenic differentiation and mitochondrial biogenesis by activating myogenin and PGC-1α in a mouse myoblast C2C12 cell line. Biochemistry and biophysics reports, 31, 101309. https://doi.org/10.1016/j.bbrep.2022.101309
Olsen, M. L., Copeland, C. E., Sundberg, C. A., Aw, R., Shaver, Z. M., Rao, G., Swartz, J. R., Karim, A. S., & Jewett, M. C. (2026). Design-driven optimization of low-cost reagent formulations for reproducible and high-yielding cell-free gene expression. Nature Communications, 17(1). https://doi.org/10.1038/s41467-026-69605-8
Available PDF from the preprint article:
Panah, B. Y., Wochner, A., Roos, T., Funkner, A., Kunze, M., & Ag, C. (2015, 23 diciembre). WO2017109161A1 - Method of rna in vitro transcription using a buffer containing a dicarboxylic acid or tricarboxylic acid or a salt thereof - Google Patents. https://patents.google.com/patent/WO2017109161A1/en
Romantseva, E., Alperovich, N., Ross, D., Lund, S. P., & Strychalski, E. A. (2022). Effects of DNA template preparation on variability in cell-free protein production. Synthetic Biology, 7(1), ysac015. https://doi.org/10.1093/synbio/ysac015
Ruiz, D. G., Sandoval-Perez, A., Rangarajan, A. V., Gunderson, E. L., & Jacobson, M. P. (2022). Cysteine oxidation in proteins: Structure, biophysics, and simulation. Biochemistry, 61(20), 2165–2176. https://doi.org/10.1021/acs.biochem.2c00349
Sato, G., Miyazawa, S., Doi, N., & Fujiwara, K. (2024). Cell-Free Protein Expression by a Reconstituted Transcription-Translation System Energized by Sugar Catabolism. Molecules (Basel, Switzerland), 29(13), 2956. https://doi.org/10.3390/molecules29132956
Siddiquee, R., & Kwan, A. (2021). Cell-free Synthesis of Correctly Folded Proteins with Multiple Disulphide Bonds: Production of Fungal Hydrophobins. BIO-PROTOCOL, 11(10), e4019. https://doi.org/10.21769/bioprotoc.4019
Silverman, A. D., Karim, A. S., & Jewett, M. C. (2020). Cell-free gene expression: an expanded repertoire of applications. Nature reviews. Genetics, 21(3), 151–170. https://doi.org/10.1038/s41576-019-0186-3
Available PDF from the article:
Cellfree gene expression an expanded repertoire of applications 2020.pdf
Sun, Z. Z., Chiao, A. C., Robertson, D. E., Metzger, L. E., IV, Mansfield, R., Trego, K. S., & Inc, S. (2023). US20200181670A1 - Improved In Vitro Transcription/Translation (TXTL) system and use thereof - Google Patents. https://patents.google.com/patent/US20200181670A1/en
Thornton, E. L., Boyle, J. T., Laohakunakorn, N., & Regan, L. (2025). Cell-Free protein synthesis as a method to rapidly screen Machine Learning-Generated protease variants. ACS Synthetic Biology, 14(5), 1710–1718. https://doi.org/10.1021/acssynbio.5c00062
Waller, A. (2024). Nuclease free water - its importance in biological research. Norgen Biotek Corp. https://norgenbiotek.com/blog/importance-water-quality-biological-research?srsltid=AfmBOoofzG840N-ZbWhRUi2ZdwS5nmivMS6KNofuDH0W2w6vVPHzF69O
Wang, H., He, X., Li, Z., Jin, H., Wang, X., & Li, L. (2022). Guanosine primes acute myeloid leukemia for differentiation via guanine nucleotide salvage synthesis. https://pmc.ncbi.nlm.nih.gov/articles/PMC8822274/
Worst, E. G., Exner, M. P., De Simone, A., Schenkelberger, M., Noireaux, V., Budisa, N., & Ott, A. (2016). Residue-specific Incorporation of Noncanonical Amino Acids into Model Proteins Using an Escherichia coli Cell-free Transcription-translation System. Journal of Visualized Experiments, 114. https://doi.org/10.3791/54273
Yadav, S., Perkins, A. J. P., Liyanagedera, S. B. W., Bougas, A., & Laohakunakorn, N. (2025). ATP Regeneration from Pyruvate in the PURE System. ACS synthetic biology, 14(1), 247–256. https://doi.org/10.1021/acssynbio.4c00697
Yusri, K., Jose, S., Vermeulen, K. S., Tan, T. C. M., & Sorrentino, V. (2025). The role of NAD+ metabolism and its modulation of mitochondria in aging and disease. Npj Metabolic Health And Disease, 3(1), 26. https://doi.org/10.1038/s44324-025-00067-0
Zhang, Y., Huang, Q., Deng, Z., Xu, Y., & Liu, T. (2018). Enhancing the efficiency of cell-free protein synthesis system by systematic titration of transcription and translation components. Biochemical Engineering Journal, 138, 47-53. https://doi.org/10.1016/j.bej.2018.07.001 [No Open Access]
Part C:
Balleza, E., Kim, J. M., & Cluzel, P. (2018). Systematic characterization of maturation time of fluorescent proteins in living cells. Nature methods, 15(1), 47–51. https://doi.org/10.1038/nmeth.4509
Botman, D., De Groot, D. H., Schmidt, P., Goedhart, J., & Teusink, B. (2019). In vivo characterisation of fluorescent proteins in budding yeast. Scientific Reports, 9(1), 2234. https://doi.org/10.1038/s41598-019-38913-z
Goedhart, J., Von Stetten, D., Noirclerc-Savoye, M., Lelimousin, M., Joosen, L., Hink, M. A., Van Weeren, L., Gadella, T. W., & Royant, A. (2012). Structure-guided evolution of cyan fluorescent proteins towards a quantum yield of 93%. Nature Communications, 3(1), 751. https://doi.org/10.1038/ncomms1738
Kremers, G., Hazelwood, K. L., Murphy, C. S., Davidson, M. W., & Piston, D. W. (2009). Photoconversion in orange and red fluorescent proteins. Nature Methods, 6(5), 355–358. https://doi.org/10.1038/nmeth.1319
Papadaki, S., Wang, X., Wang, Y., Zhang, H., Jia, S., Liu, S., Yang, M., Zhang, D., Jia, J., Köster, R. W., Namikawa, K., & Piatkevich, K. D. (2022). Dual-expression system for blue fluorescent protein optimization. Scientific Reports, 12(1), 10190. https://doi.org/10.1038/s41598-022-13214-0
Pédelacq, J. D., Cabantous, S., Tran, T., Terwilliger, T. C., & Waldo, G. S. (2006). Engineering and characterization of a superfolder green fluorescent protein. Nature biotechnology, 24(1), 79–88. https://doi.org/10.1038/nbt1172
Pharma, B. (2025, November 25). SFGFP – Recombinant Fluorescent Protein. Basic Pharma. https://basicpharma.nl/biotech/recombinant-protein-supplier/sfgfp
Part D:
Bioworks, G. (2026). Ginkgo Bioworks Launches Ginkgo Cloud Lab, Powered by Autonomous Lab Infrastructure. PR NEWSWIRE. [https://www.prnewswire.com/news-releases/ginkgo-bioworks-launches-ginkgo-cloud-lab-powered-by-autonomous-lab-infrastructure-302700458.html(https://www.prnewswire.com/news-releases/ginkgo-bioworks-launches-ginkgo-cloud-lab-powered-by-autonomous-lab-infrastructure-302700458.html)]
Ginkgo Bioworks. (2026). Automation. https://www.ginkgo.bio/automation-product
Wintermute, J. (2024, September 12). Why reconfigurable lab automation sparks joy. Foundry Theory. https://foundrytheory.substack.com/p/reconfigurable-lab-joy
Sources:
Part C.
| Fluorescent Protein | Webpage |
|---|---|
| sfGFP | https://www.fpbase.org/protein/superfolder-gfp/ |
| mRFP1 | https://www.fpbase.org/protein/mrfp1/ |
| mKO2 | https://www.fpbase.org/protein/mko2/ |
| mTurquoise2 | https://www.fpbase.org/protein/mturquoise2/ |
| mScarlet_I | https://www.fpbase.org/protein/mscarlet/ |
| Electra2 | https://www.fpbase.org/protein/electra2/ |
Part D.
Assembly Output from: https://racs.rcdonovan.com/
| Model | In assemble characteristics |
|---|---|
| (rac-E17) | Z rot: 90°, Instrument: Generic 0.3×0.1×0.1, Color RGB: 0, 251, 255 |
| mm-1m | Z rot: 0° |
| Lunatic (rac-E15) | Z rot: 270°, Instrument: Lunatic, Generic 0.22×0.15×0.12, Color RGB: 254, 6, 208 |
| mm-1m | Z rot: 90° |
| (rac-E15) | Z rot: 270°, Instrument: Generic 0.4×0.1×0.1, Color RGB: 255, 15, 191 |
| Lunatic (rac-E15) | Z rot: 270°, Instrument: Lunatic |
| mm-1m | Z rot: 0° |
| taxi-5puck | Z rot: 0° |
| rac-E16 | Z rot: 0° |
| rac-E16 | Z rot: 0° |
| rac-E16 | Z rot: 0° |
| mm-switch | Z rot: 180° |
| mm-switch | Z rot: 180°, flip horizontal |
| mm-1m | Z rot: 0° |
| (rac-E17) | Z rot: 0°, Instrument: Generic 0.45×0.26×0.2, Color RGB: 255, 0, 51 |