Individual Final Project
A. Proposal Ideas

Idea 1: Kinetic Bio-Interface
Sources of Inspiration: MPU-6050 IMU Sensor + Electrobacteria (Shewanella/Geobacter) + Pattern Recognition
Problem/Vision: It is easy to translate human movement into digital systems, but translating it into a “living/biological” system is very difficult. Can we design a device that bridges human kinetics and microbial metabolism?
Mechanism: A wearable MPU-6050 accelerometer sensor (IMU) reads the user’s hand or dance movements (gesture recognition). Machine learning algorithms analyze these movement patterns (pattern recognition) and convert them into small electrical signals. These signals are transmitted to electro-bacteria such as Shewanella oneidensis or Geobacter living on a microfluidic chip. The electro-bacteria take in this electron flow from the electrodes (microbial electrosynthesis), alter their metabolism, and produce a movement-specific response (e.g., a color change or fluorescent emission).
Automation (Opentrons): The robot is used to distribute different bacterial concentrations and conductive liquid media onto 96-well plates to test for the ideal “kinetic biosolution” where the electrical signal is transmitted most effectively.
Simulation
Strengths (+)
Interdisciplinary depth! Wearable sensor technology, machine learning, microfluidics, and synthetic biology all come together in a single project. Shewanella and Geobacter are organisms with well-characterized electron transfer mechanisms—in other words, we’re not inventing things from scratch; we’re building upon existing biology. Integrating with Opentrons also moves the project from a “conceptual demo” to “systematic optimization,” which is highly valuable.
Challenges / Risks (-)
The signal-bacteria interface is the most critical bottleneck. Electro-bacteria operate at microampere-level currents, and aligning signals derived from IMU data with this threshold requires significant calibration. Real-time response is also challenging: bacterial metabolism responds on the order of minutes, not seconds, so the “instant dance → luminescence” scenario will actually require a delay buffer. Microfluidic chip design and fabrication is a project in itself.
Decision on development feasibility: It can be developed, but with a narrowed scope. Perhaps simply getting the “single gesture → single bacterial response” loop to work for the final demo would be a very powerful demonstration. Attempting to run the full pipeline carries the risk of not being able to demonstrate any single layer in depth.
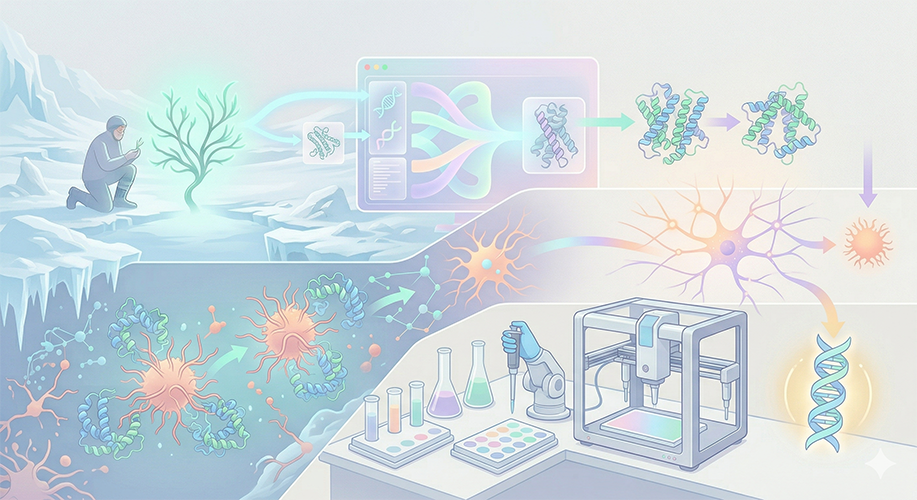
Idea 2: Paleo-Proteins (Neurological Protection Through Ice Age Resilience)
Waving to my Archeology background which I couldn’t complete 🫣 (Double Major)
Sources of Inspiration: Revival of plants from the Ice Age + Synthetic Antibody/Protein Design Using LLM + The Link Between Alzheimer’s and Dementia and Proteins
Problem/Vision: At the root of Alzheimer’s and dementia lies the accumulation of misfolded proteins in the brain. Ancient organisms that have remained dormant since the Ice Age and can revive possess remarkable chaperone (protective) protein mechanisms that prevent their proteins from freezing and breaking down.
Mechanism: Using Large Language Models (LLMs) and AI-based protein design tools (ProteinMPNN, ESMFold), we extract “language” from the protein structures of organisms that survived the Ice Age. With this AI model, we generate “Ancient Resilient Synthetic Chaperones” (Paleo-Proteins) that have the potential to prevent misfolding in Alzheimer’s plaques.
Automation (Opentrons): Dozens of different synthetic protein variants generated by AI are produced in cell-free systems. The Opentrons robot automatically screens these proteins by mixing them with target (amyloid/tau-like) proteins to determine which variant best prevents aggregation/folding (High-Throughput Screening).
Simulation
Strengths (+)
Narrative power! The statement “Ice Age organisms could treat Alzheimer’s” is both scientifically defensible and a compelling bio-narrative. My background in archaeology combined with biology perfectly aligns with the HTGAA spirit: I am the prime example of the archetype of someone who comes from a different discipline and views biotechnology differently.
It’s technically sound as well: chaperone proteins from permafrost organisms are well-characterized; ProteinMPNN and ESMFold are production-ready tools; cell-free protein expression with Opentrons is highly suitable for automation; and the ThT fluorescence assay is a standard method for measuring amyloid. Every layer is grounded in existing technologies.
Risks (-)
The biggest risk is the massive translation gap between synthetic chaperones preventing amyloid aggregation in vitro and neuroprotection in vivo; but this is a final project, not a clinical trial, so an in vitro demonstration is more than sufficient. Protein folding quality must also be controlled in cell-free expression; a misfolded chaperone would be ironic.
Paleo-Proteins has a much clearer “problem → solution” pathway compared to my other suggestions. The Kinetic Bio-Interface is creative, but the “what’s the point” question remains unclear; here, the target is Alzheimer’s, the mechanism of action is clear, and the measurement method is standard. (Every project should also consider the sponsor’s perspective, and this idea is in a position to attract more support.)
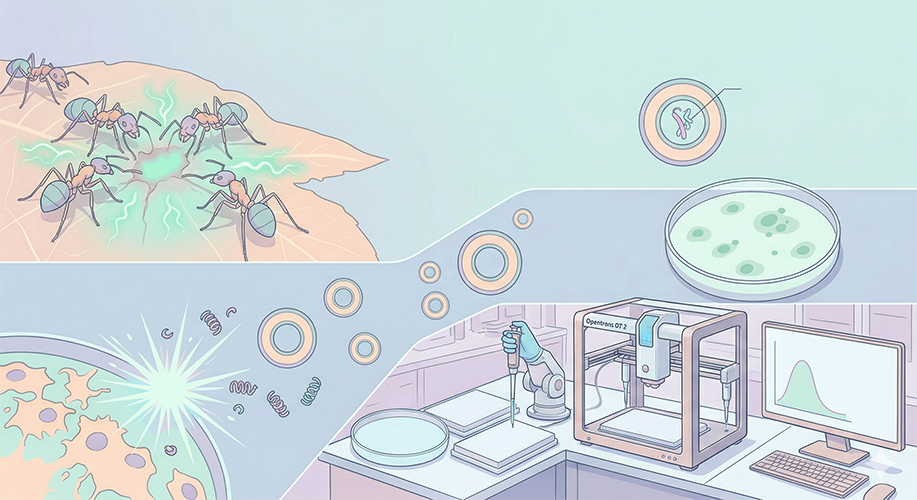
Idea 3: Bio-Swarm (Ant-Algorithm Micro-Surgical Droplets)
Sources of Inspiration: Surgeon ants + Microfluidics + Pattern Recognition
Problem/Vision: In nature, certain ant species recognize the infection scent/chemical (pattern) in the wounds of injured colony members and secrete specific antibiotics or perform bio-surgery on that area. Current drugs spread throughout the entire body. Can we build an ant-like system that delivers the drug only to the “problematic area”?
Mechanism: Using microfluidic technology, we produce liposomes (extracellular artificial cells) embedded with synthetic genetic circuits. These artificial droplets are designed to open only when they recognize a specific disease/infection chemical pattern (just like surgeon ants) and secrete their healing (surgical/antimicrobial) protein locally.
Automation (Opentrons): Opentrons drips target “infection chemicals” onto a petri dish or microfluidic chip in a maze-like pattern. It then sets up an automated experimental apparatus to visualize how accurately the synthetic surgical droplets detect and burst (respond to) these targets.
Simulation
Strengths (+)
Narratively, it’s very striking! The “surgeon ants” metaphor is memorable, and drawing inspiration from nature (bio-inspired design) is an approach HTGAA really loves. Liposome-based targeted drug delivery is already an active area of research, but combining it with synthetic genetic circuits offers a fresh perspective. The maze test on a microfluidic chip has the potential to be a visually striking demo compatible with Opentrons.
Risks (-)
This is the most technically challenging project idea. It requires integrating three separate systems, each complex in its own right: synthetic genetic circuit design (which is a project in itself), liposome fabrication, and microfluidic navigation behavior on top of these. The “ant algorithm” part is a nice metaphor, but in reality, liposomes performing active chemotaxis is a very different engineering challenge from passive diffusion — to achieve this, receptors must be placed on the liposome surface, which requires a deep understanding of protein engineering.
Additionally, the “trigger mechanism that bursts and releases the drug” (e.g., pH-sensitive or enzyme-sensitive liposomes) and “specific chemical pattern recognition” are both highly complex when considered together. Instead of demonstrating all of these in the final demo, it would be better to focus on just one aspect — such as the “opening of liposomes sensitive to infection markers” — and present that in depth.
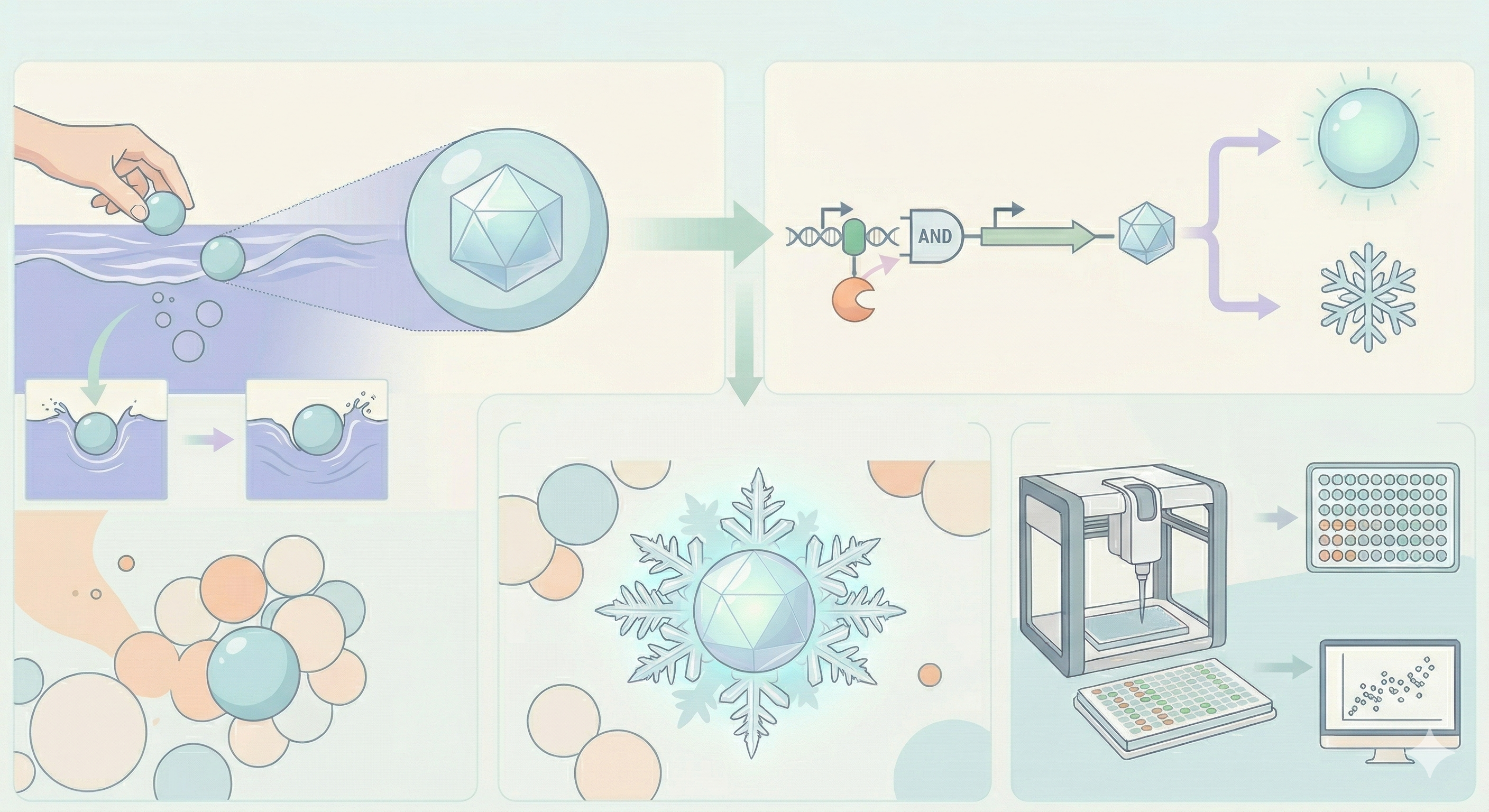
Idea 4: Cryo-Lume
Spherified Biosensors for Aesthetic Contaminant Detection via Halochromic and Cryogenic Phase-Change
Background
Current environmental biosensors predominantly rely on simple colorimetric or fluorescent readouts to detect contaminants. Cryo-Lume proposes a paradigm shift in bio-art and bioremediation by introducing a dual-output biosensor that couples bioluminescence with a macroscopic physical phase change—specifically, localized ice nucleation in a supercooled fluid.
The biological chassis will be encapsulated within calcium alginate spheres. While sodium alginate is naturally highly hydrophilic, calcium alginate can be engineered to be hydrophobic for structural and bioremediation applications. The physical behavior of these hydrophobic spheres when introduced into an aquatic environment is critical; their specific water-entry dynamics and fluid displacement determine how effectively they interface with the sample fluid (Watson et al., 2025).
Upon entering the contaminated water, the engineered bacteria inside the hydrophobic spheres will detect specific pollutants (e.g., heavy metals/arsenic) and simultaneously activate two distinct pathways: the luxCDABE operon for autonomous bioluminescence, and the inaK or inaZ gene for ice nucleation. This causes the surrounding supercooled water to rapidly freeze around the sphere, forming a glowing, precipitating “snowflake” that visually and physically captures the contaminant.
Aim
Design and Characterize the Dual-Output Genetic Circuit. Construct an AND-gate logic circuit combining a contaminant-responsive promoter (e.g., ArsR) with the lux operon (for glowing) and the inaK protein (for ice nucleation).
Optimize the Hydrophobic Spherification Process. Produce calcium alginate capsules containing the engineered cells, modifying the polymer to achieve the specific hydrophobic water-entry dynamics and fluid interaction parameters outlined by Watson et al. (2025).
Automate Biosensor Screening and Calibration. Utilize the Opentrons liquid handling robot to automate reaction setups in 96-well plates. This high-throughput screening will allow us to test various alginate concentrations and characterize the signal-to-noise ratio, mapping exact contaminant levels to their corresponding cryogenic/luminescent response times.
Simulation
Strengths (+)
The dual-output design is genuinely novel. Existing biosensors produce a single readout (color, fluorescence, or electrochemical signal) that requires instrumentation to read. Cryo-Lume produces two simultaneous outputs: a molecular signal (bioluminescence, quantitative via luminometry) and a macroscopic physical phase change (ice nucleation, visible to the naked eye). This means the biosensor works both in a lab with a plate reader AND in a remote village with no equipment — you literally see it glow and feel the ice form. Both biological components — luxCDABE and inaK — are among the most well-characterized systems in synthetic biology. The lux operon has been used in biosensors since the 1990s with published dose-response curves. The inaK ice nucleation protein from Pseudomonas syringae has been studied for decades in atmospheric science and food technology. Neither requires speculative biology — this is engineering with proven parts. The real-world problem is enormous and urgent: arsenic contamination affects over 200 million people globally, predominantly in communities with the least access to analytical infrastructure. A low-cost, equipment-free biosensor directly addresses environmental justice.
Challenges / Risks (-)
Maintaining supercooled water at -3°C requires careful temperature control, though the ~8°C gap between inaK-induced and spontaneous nucleation provides a comfortable margin. The hydrophobic alginate coating could slow arsenic diffusion if too thick, but this is precisely what the Opentrons optimization matrix is designed to calibrate. Cell viability in spheres degrades over time — fine for a final demo with fresh preparations, but long-term deployment would need freeze-drying. All risks have clear mitigations and none are fundamental blockers.
B. FINAL PROPOSAL
Paleo-Proteins
Synthetic Cryoprotectants for Therapeutic Hypothermia and Tissue Preservation
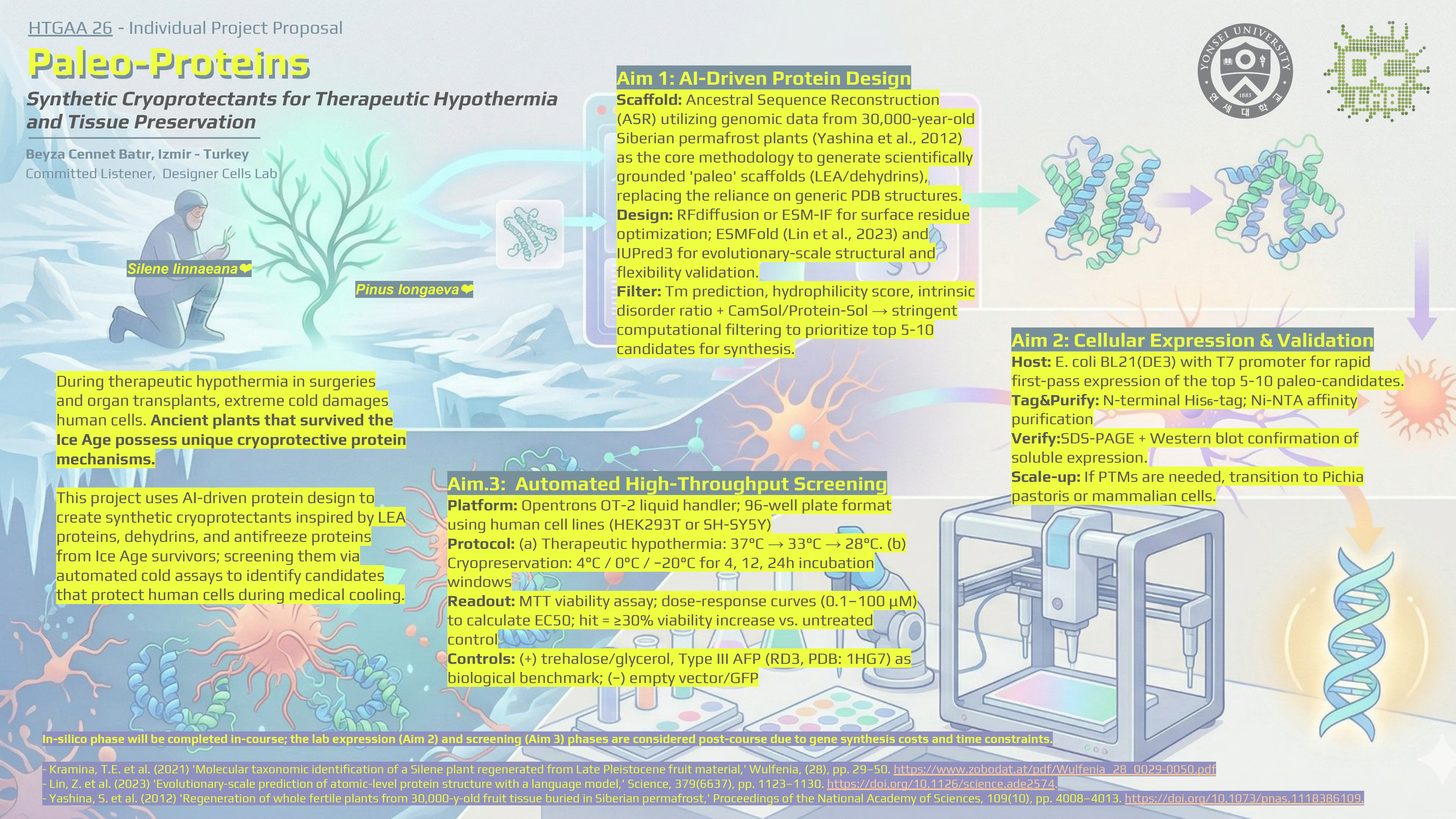
During therapeutic hypothermia in surgeries and organ transplants, extreme cold damages human cells. Ancient plants that survived the Ice Age possess unique cryoprotective protein mechanisms.
This project uses AI-driven protein design to create synthetic cryoprotectants inspired by LEA proteins, dehydrins, and antifreeze proteins from Ice Age survivors; screening them via automated cold assays to identify candidates that protect human cells during medical cooling.
Aim 1: AI-Driven Protein Design
- Scaffold: Ancestral Sequence Reconstruction (ASR) utilizing genomic data from 30,000-year-old Siberian permafrost plants (Yashina et al., 2012) as the core methodology to generate scientifically grounded ‘paleo’ scaffolds (LEA/dehydrins), replacing the reliance on generic PDB structures.
- Design: RFdiffusion or ESM-IF for surface residue optimization; ESMFold (Lin et al., 2023) and IUPred3 for evolutionary-scale structural and flexibility validation.
- Filter: Tm prediction, hydrophilicity score, intrinsic disorder ratio + CamSol/Protein-Sol → stringent computational filtering to prioritize top 5-10 candidates for synthesis.
Aim 2: Cellular Expression & Validation
- Host: E. coli BL21(DE3) with T7 promoter for rapid first-pass expression of the top 5-10 paleo-candidates.
- Tag&Purify: N-terminal His₆-tag; Ni-NTA affinity purification
- Verify:SDS-PAGE + Western blot confirmation of soluble expression.
- Scale-up: If PTMs are needed, transition to Pichia pastoris or mammalian cells.
Aim.3: Automated High-Throughput Screening
- Platform: Opentrons OT-2 liquid handler; 96-well plate format using human cell lines (HEK293T or SH-SY5Y)
- Protocol: (a) Therapeutic hypothermia: 37°C → 33°C → 28°C. (b) Cryopreservation: 4°C / 0°C / −20°C for 4, 12, 24h incubation windows
- Readout: MTT viability assay; dose-response curves (0.1–100 μM) to calculate EC50; hit = ≥30% viability increase vs. untreated control
- Controls: (+) trehalose/glycerol, Type III AFP (RD3, PDB: 1HG7) as biological benchmark; (−) empty vector/GFP
We are not just looking at modern cold-hardy plants. The ‘Paleo-Proteins’ pipeline targets the genomic signatures of 30,000-year-old plant tissues buried in Siberian permafrost (Yashina et al., 2012). Although initially misidentified by morphology, recent molecular phylogenetic analyses have confirmed that these ancient survivors do indeed belong to the Silene linnaeana group (Kramina et al., 2021). This firm confidence in molecular truth rather than pure morphology is also the reason for using Ancestral Sequence Reconstruction (ASR) combined with evolutionary scale models such as ESMFold in Goal 1 (Lin et al., 2023).
Simulation
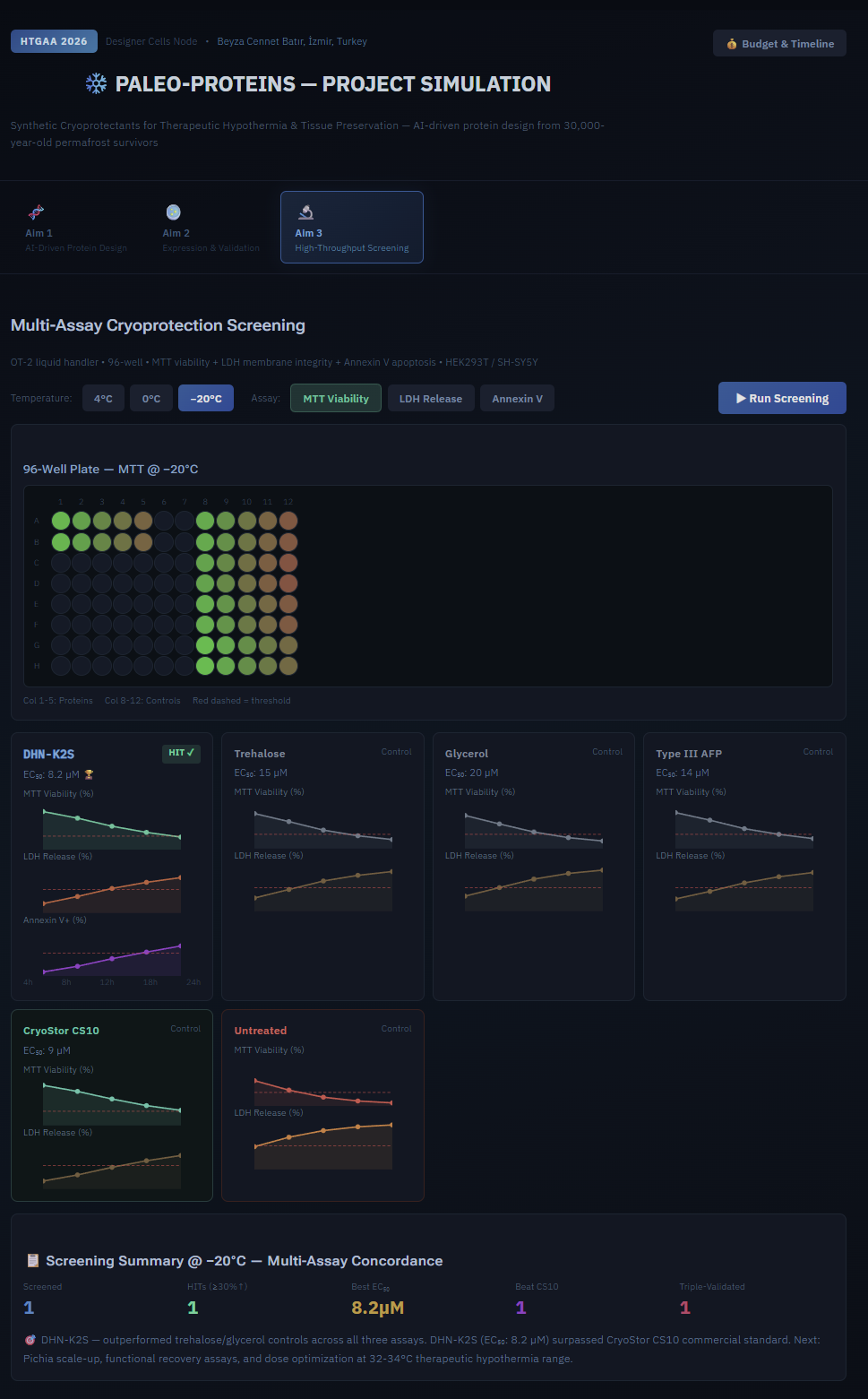
- According to simulation data, DHN-K2S (EC₅₀: 8.2 µM) is the most promising candidate—it passes all tests.
In-silico phase will be completed in-course; the lab expression (Aim 2) and screening (Aim 3) phases are considered post-course due to gene synthesis costs and time constraints.
Situated at the intersection of synthetic biology and quantum thermodynamics, Paleo-Proteins reimagines cellular cryopreservation, drawing inspiration from the physics of ultracold matter. Just as a single ‘impurity’ or ‘polaron’ can dynamically alter the energy, size, and motion of an ultracold Bose-Einstein condensate, AI-designed synthetic chaperones act as protective ‘impurities’ within the frozen cytoplasm, thermodynamically disrupt ice crystallization, and preserve cellular integrity (Simons, 2022). To validate these synthetic cryoprotectants, we will apply the meticulous calibration, control, and precise measurement practices inherent to quantum physics to Opentrons’ automated high-throughput screening, enabling the extraction of accurate signals from these noisy biological cell viability data. Ultimately, this project—which integrates AI-driven protein design, medical science, and the dynamics of ultra-cold fluids—aims to engineer unprecedented biological resilience during therapeutic hypothermia and push the boundaries of human lifespan through tissue preservation (Simons, 2025).
Organizations of Interest
Twist Biosciences: Whole plasmid synthesis for all Paleo-Protein expression constructs Ginkgo Bioworks: Primary lab automation (Echo525, PHERAstar FSX, Cytomat) and CFPS master mix Asimov Kernel: DNA construct design, circuit simulation, and construct registry Opentrons: OT-2 automated liquid handling for cell seeding and MTT assay workflow Thermo Fisher Scientific: Cell culture reagents, cDNA synthesis kit, BCA assay, labware Millipore Sigma: MTT reagent, IPTG, anti-His₆ antibody, trehalose positive control New England Biolabs: BL21(DE3) competent cells for recombinant protein expression SecureDNA: Biosecurity screening of all synthetic DNA orders
Future: DeepCure, Takeda Pharmaceuticals
References
Bikard, D., Euler, C. W., Jiang, W., Nussenzweig, P. M., Goldberg, G. W., Duportet, X., Fischetti, V. A. and Marraffini, L. A. (2014) ‘Exploiting CRISPR-Cas nucleases to produce sequence-specific antimicrobials’, Nature Biotechnology, 32, pp. 1146–1150.
Carbon Minds (n.d.) DNA as a Testament to Humanity: Transmuting social inequality indices and human rights principles into genetic code and storing them in Bacillus subtilis spores for millennia via cryptobiosis. Guillermo Romero Tecua.
Citorik, R. J., Mimee, M. and Lu, T. K. (2014) ‘Sequence-specific antimicrobials using efficiently delivered RNA-guided nucleases’, Nature Biotechnology, 32, pp. 1141–1145.
Dauparas, J. et al. (2022) ‘Robust deep learning–based protein sequence design using ProteinMPNN’, Science, 378(6615), pp. 49–56. DOI: 10.1126/science.add2187
Eriksson, S.K. et al. (2011). Phosphorylation of the dehydrin Lti29 in Arabidopsis thaliana is associated with cold acclimation. Plant, Cell & Environment, 34(7), 1071–1082. https://doi.org/10.1111/j.1365-3040.2011.02304.x
Frank, E. T., Kesner, L., Liberti, J. et al. (2023) ‘Targeted treatment of injured nestmates with antimicrobial compounds in an ant society’, Nature Communications, 14, 8446. Available at: https://doi.org/10.1038/s41467-023-43885-w
Jewett, M.C. and Swartz, J.R. (2004) ‘Mimicking the Escherichia coli cytoplasmic environment activates long‐lived and efficient cell‐free protein synthesis,’ Biotechnology and Bioengineering, 86(1), pp. 19–26. https://doi.org/10.1002/bit.20026.
Kramina, T.E. et al. (2021) ‘Molecular taxonomic identification of a Silene plant regenerated from Late Pleistocene fruit material,’ Wulfenia, (28), pp. 29–50. https://www.zobodat.at/pdf/Wulfenia_28_0029-0050.pdf
Lin, Z. et al. (2023) ‘Evolutionary-scale prediction of atomic-level protein structure with a language model,’ Science, 379(6637), pp. 1123–1130. https://doi.org/10.1126/science.ade2574.
Meighen, E.A. (1991) ‘Molecular biology of bacterial bioluminescence,’ Microbiological Reviews, 55(1), pp. 123–142. https://doi.org/10.1128/mr.55.1.123-142.1991.
Min, J., Kim, E. J., LaRossa, R. A. and Gu, M. B. (2000) ‘Detection of environmental effects using recombinant bioluminescent Escherichia coli strains’, Radiation and Environmental Biophysics, 39(1), pp. 41–45.
New understanding of why supercooled water droplets sometimes explode when they freeze’ (2023) Chemistry World. Available at: https://www.chemistryworld.com/news/new-understanding-of-why-supercooled-water-droplets-sometimes-explode-when-they-freeze/4017904.article (Accessed: 3 April 2026).
Oxford University Cryogenic Fluid Dynamics Lab (n.d.) Research on multiphase cryogenic processes, liquid hydrogen, and cryogenic carbon capture. Available at: http://eng.ox.ac.uk/cryogenic-fluid-dynamics-lab
Sarkar, K., Bonnerjee, D. and Bagh, S. (2021) ‘Distributed computing with engineered bacteria and its application in solving chemically generated 2×2 maze problems’, ACS Synthetic Biology, 10(10), pp. 2456–2464. Available at: https://pubs.acs.org/doi/10.1021/acssynbio.1c00279
Schaefer, V. J. (1946) ‘The production of ice crystals in a cloud of supercooled water droplets’, Science, 104(2707), pp. 457–459. Available at: https://doi.org/10.1126/science.104.2707.457
‘Scientists Discover Water That’s Frozen and Liquid at Once’ (2025) ScienceBlog.com. Available at: https://scienceblog.com/scientists-discover-water-thats-frozen-and-liquid-at-once/ (Accessed: 3 April 2026).
Shi, L. et al. (2016) ‘Extracellular electron transfer mechanisms between microorganisms and minerals,’ Nature Reviews Microbiology, 14(10), pp. 651–662. https://doi.org/10.1038/nrmicro.2016.93.
Simons, L. (2025). Bose polarons in superfluids and supersolids [Doctoral dissertation, University of Antwerp] https://repository.uantwerpen.be/docman/irua/d8fd0bmotoM35
Simons, L. (2021). Analog physics with excitations in Bose-Einstein Condensates [Master’s thesis, Universiteit Antwerpen] https://medialibrary.uantwerpen.be/files/7606/704c6cf3-5f9c-4a15-989f-98fed6ffd3b2.pdf
Soprunyuk, V. and Schranz, W. (2023) ‘Glass and freezing transition of supercooled water confined in mesoporous materials and biological systems,’ International Online Medical Council [Preprint]. https://doi.org/10.35248/2322-3308-12.4.001.
Tunnacliffe, A. and Wise, M.J. (2007) The continuing conundrum of the LEA proteins, Naturwissenschaften. journal-article, pp. 791–812. https://doi.org/10.1007/s00114-007-0254-y.
Watson, D.A. et al. (2025) ‘Water entry dynamics of hydrophobic spheres bounded by semi-infinite cylindrical pipes,’ Physics of Fluids, 37(11). https://doi.org/10.1063/5.0300541.
‘Wilson Bentley (Snowflake Bentley)’ (n.d.) Wikipedia. Available at: https://en.wikipedia.org/wiki/Wilson_Bentley
Wogan, T. (2024) ‘Experimental proof of liquid–liquid transition in supercooled water’, Chemistry World. Available at: https://www.chemistryworld.com/news/experimental-proof-of-liquid-liquid-transition-in-supercooled-water/4012820.article
Wu, J. and Rosen, B.P. (1993) ‘Metalloregulated expression of the ars operon.,’ Journal of Biological Chemistry, 268(1), pp. 52–58. https://doi.org/10.1016/s0021-9258(18)54113-2.
Yashina, S., Gubin, S., Maksimovich, S., Yashina, A., Gakhova, E. and Gilichinsky, D. (2012) ‘Regeneration of whole fertile plants from 30,000-y-old fruit tissue buried in Siberian permafrost’, Proceedings of the National Academy of Sciences, 109(10), pp. 4008–4013. Available at: https://doi.org/10.1073/pnas.1118386109
I used Claude for all technical infrastructure and simulation testing and generated the visuals using NanoBanana 2.
I would like to thank our Node Chair Prof. Han, for his valuable contributions and feedback which helped me refine the final project idea.
C. HTGAA 2026: Individual Final Project Documentation
SECTION 1: ABSTRACT
1. Abstract
Therapeutic hypothermia and cryopreservation are critical procedures in medical surgeries, organ transplants, and cellular therapies. However, extreme cold temperatures inherently cause cellular damage, crystallization, and tissue degradation. At the root of this challenge is the need for effective cryoprotectants that can thermodynamically disrupt ice crystallization and preserve cellular integrity. To address this, the “Paleo-Proteins” project draws inspiration from evolutionary biology—specifically, ancient plants that survived the Ice Age, such as the Silene linnaeana group found in 30,000-year-old Siberian permafrost. These ancient organisms possess unique, highly resilient chaperone protein mechanisms.
The broad objective of this project is to harness AI-driven protein design to develop synthetic cryoprotectants inspired by Late Pleistocene Late Embryogenesis Abundant (LEA) proteins and dehydrins. We hypothesize that computationally generated “paleo” scaffolds, optimized via modern AI models, will act as protective ‘impurities’ within the frozen cytoplasm and exhibit superior cryoprotective properties compared to generic proteins. The specific aims involve utilizing Ancestral Sequence Reconstruction (ASR) to generate scaffolds, optimizing them with RFdiffusion and ESM-IF, and filtering candidates using ESMFold and IUPred3.
Ultimately, this project aims to express these candidates and validate their efficacy via automated high-throughput cold assays, establishing a novel pipeline for engineering unprecedented biological resilience during therapeutic cooling.
SECTION 2: PROJECT AIMS
AIM.1: Experimental Aim The first aim of my final project is to computationally design a library of ‘Ancient Resilient Synthetic Chaperones’ (Paleo-Proteins) by utilizing Ancestral Sequence Reconstruction (ASR) and AI-based protein design tools. Relevant methods and resources:
Ancestral Sequence Reconstruction (ASR): Mining and reconstructing genomic data from 30,000-year-old Ice Age permafrost survivors (Silene linnaeana).
AI Protein Design Tools: Utilizing RFdiffusion or ESM-IF for de novo surface residue optimization, alongside ESMFold and IUPred3 for atomic-level 3D structural prediction and flexibility validation.
Computational Filtering: Applying computational models to screen candidates based on melting temperature (Tm) predictions, hydrophilicity scores, and intrinsic disorder ratios (CamSol/Protein-Sol) to prioritize the top 5-10 candidates.
AIM.2: Experimental Aim
The next step following a successful in-silico design (Aim 1) would be to transition into physical synthesis and automated in-vitro testing. This involves synthesizing the DNA sequences of the top AI-generated protein candidates via Twist Bioscience and expressing them in model cell lines (e.g., E. coli BL21).
To address the technical challenge of high-throughput testing, an Opentrons OT-2 liquid-handling robot will be deployed to automate the screening workflow. The robot will format 96-well plates, subject the engineered cells to a gradient of sub-body/freezing temperatures (simulating therapeutic hypothermia), and automate MTT cell viability assays to pinpoint which specific variants successfully confer cold resistance.
AIM.3: Visionary Aim
The long-term vision for this project is to translate these synthetic cryoprotectants into clinical therapeutics, directly addressing a major barrier in the medical field: the inherent cellular damage caused by extreme cold during therapeutic hypothermia.
If fully realized, these “Paleo-Proteins” would challenge the existing paradigm of tissue preservation by acting as protective ‘impurities’ (conceptually akin to polarons stabilizing ultracold quantum matter) that thermodynamically disrupt ice crystallization inside human cells.
This would revolutionize clinical practices during complex surgeries, organ transplants, and brain trauma management, safely extending the viability window for human organs and ultimately pushing the boundaries of human longevity.
SECTION 3: BACKGROUND
3a. Background and Literature Context
Therapeutic hypothermia is a critical clinical practice used to preserve organ function and minimize brain trauma during complex surgeries, yet the inherent cellular damage caused by extreme cold remains a major barrier.
Current cryopreservation methods lack targeted, highly efficient biological protectants that operate effectively within human tissue at sub-body temperatures without causing toxicity. To address this gap, this project draws core biological inspiration from Yashina et al. (2012), who successfully regenerated whole fertile plants from 30,000-year-old fruit tissue buried in Siberian permafrost, demonstrating the remarkable, long-term viability of ancient cryoprotective mechanisms.
Complementarily, the physical principles of this project are inspired by Simons (2025), whose research on Bose polarons in ultracold matter illustrates how single “impurities” can dynamically alter the energy, size, and motion of supercooled environments. By integrating these biological and physical insights, we aim to design synthetic chaperones that act as protective ‘impurities’ within freezing cytoplasm to prevent crystallization.
3b. How the project is novel or innovative
Unlike traditional drug discovery that relies on existing modern protein structures, this project utilizes Ancestral Sequence Reconstruction (ASR) combined with evolutionary-scale AI models (like ESMFold) to mine and revive ancient genomic data from Ice Age survivors.
Furthermore, it reconceptualizes biological freezing through the lens of quantum thermodynamics, treating the engineered synthetic chaperones as functional ‘impurities’ (akin to polarons) that thermodynamically disrupt ice crystallization inside the cell. This highly interdisciplinary approach expands the boundaries of synthetic biology by merging evolutionary archaeology, artificial intelligence, and ultracold fluid dynamics to challenge the current biological limits of cellular cold resistance.
3c. Why the project matters and what impact it could have
During complex medical procedures such as organ transplants, major cardiovascular surgeries, or brain trauma management, inducing sub-body temperatures (therapeutic hypothermia) is crucial to temporarily halt cellular decay and increase patient survival . However, a pressing real-world problem is that extreme cold inherently damages human cells, representing a critical barrier to progress in these life-saving surgical practices . This project attempts to solve this paradox by developing therapeutic “Paleo-Proteins” that act as synthetic cryoprotectants to safely prevent cold-induced cellular damage.
If successful, the broader societal contribution would be immense; it could significantly extend the viability window for donor organs—easing the global organ shortage crisis—and reduce irreversible brain damage in trauma patients . Ultimately, this advancement would drastically alter clinical practices in hypothermic medicine, improve the safety of complex surgeries, and push the boundaries of human longevity by enabling an entirely new biological capability: engineered resilience against extreme cold.
3d. Ethical implications associated with project and relevant ethical principles The development of “Paleo-Proteins” as synthetic cryoprotectants for therapeutic hypothermia holds profound implications for public health, directly aligning with the ethical principle of beneficence. By extending the viability window for donor organs and minimizing cellular damage during complex brain or cardiovascular surgeries, this research addresses a critical medical need and aims to save lives. However, because this technology fundamentally pushes the boundaries of tissue preservation and human longevity, it introduces severe implications regarding justice. As seen in emerging longevity research, such advancements raise crucial questions about equity, consent, and accessibility. If these enhanced cryoprotectants are successfully developed, there is a risk they could be restricted exclusively to high-resource medical facilities or wealthy individuals, thereby widening the global healthcare inequality gap. Furthermore, the principle of non-maleficence must be prioritized; engineering extreme cold resistance into biological expression systems (such as the E. coli used for production) presents a potential biosafety hazard if these resilient traits were to escape into the environment and disrupt local ecosystems.
To ensure the ethical execution of this project, I propose implementing strict biocontainment actions—such as engineered genetic kill-switches or synthetic amino acid auxotrophy—in our microbial chassis, alongside transparent, equitable licensing models to guarantee broad public health access to the final therapeutic. A potential unintended consequence of successfully creating highly resilient “super-cryoprotectants” is that they might inadvertently encourage unsafe, speculative procedures (such as unregulated extreme body cooling or commercial longevity biohacking) outside of approved clinical settings. Furthermore, we must acknowledge crucial scientific uncertainties: we could be wrong in our assumption that AI-designed ancient protein analogs will be safe and non-immunogenic in vivo. Medical AI models can overfit, and introducing foreign protein structures into the human bloodstream might trigger severe, unforeseen immune responses. Given these risks, a viable alternative to our proposed action of administering synthetic proteins directly to patients would be to use our in vitro automated screening platform solely to discover non-biological, small-molecule drugs that mimic the thermodynamic properties of Paleo-Proteins. This alternative would offer a cheaper, less immunogenic, and more universally accessible solution for global public health.
SECTION 4: EXPERIMENTAL DESIGN, TECHNIQUES, TOOLS, AND TECHNOLOGY
- Claude | Final Project Experimental Design
Please use this directing link to see my final project proposal which is created via Claude Code based on a skill developed by Ronan Donovan.
- Detailed Experimental Plan & Timeline
Sub-aim 1: In-Silico AI Protein Design and DNA Assembly (Weeks 1-3)
- Reminder: All HTGAA projects must include some DNA design! Make sure this form is submitted.
- Please use this directing link to see my submitted form.
- Techniques relevant to the project
- Foundational Lab Practices
- Pipetting
- Lab Safety
- Bioethical Considerations (must check this box)
- DNA Skills & Analysis
- DNA Gel Art
- DNA Sequencing
- DNA Editing (e.g., CRISPR)
- DNA Construct Design
- Restriction Enzyme Digestion
- Gel Electrophoresis
- DNA Purification from Gel
- Databases (e.g., GenBank, NCBI, Ensembl, UCSC Genome Browser)
- Laboratory Automation
- Opentrons
- Creating Code for Laboratory Automation
- Using Liquid Handling Robots (e.g., Opentrons)
- Opentrons
- Protein Design
- Protein Design
- Models and Notebooks
- Databases
- Tools
- Protein Design
- BioProduction
- [x] Chassis Selection (e.g., DH5α, Bl21-DE3 for expression)
- Registry of Standard Biological Parts
- FreeGenes
- Plasmid Preparation
- Bacterial Culturing
- Quality Control / Analysis
- Bacterial Processing (e.g., Centrifugation, Lysis, DNA Purification)
- Cell-Free System
- Cell-Free Reactions
- Freeze-Dried Cell-Free Systems
- NEB Express Kits
- miniPCR Tools
- Gibson Assembly
- Primer Design or Selection
- PCR Reactions
- Gibson Assembly
- Other Cloning Methods (e.g., Restriction Enzyme Digestion or Gateway Cloning)
- Creating Twist Order
4. Two expanded techniques by describing how I would utilize those techniques in final project.
Technique 1 - AI Protein Design: AI-driven protein design is the foundational technology of this project — without it, generating a library of cryoprotective candidates from ancient genomic data would be impossible within any realistic experimental timeline. In this project, I use a three-stage computational pipeline: RFdiffusion generates diverse protein backbone geometries conditioned on the canonical dehydrin K-segment amphipathic helix motif (EKKGIMDKIKEKLPG), exploring structural space that no natural evolutionary trajectory has visited; ESM-IF1 then performs inverse folding on each backbone, outputting amino acid sequences predicted to fold into those geometries while simultaneously satisfying evolutionary plausibility constraints learned from 250 million natural protein sequences; and ESMFold validates each candidate by predicting its full 3D structure from sequence alone, with per-residue pLDDT confidence scores used to confirm that K-segment regions adopt the expected α-helical geometry while spacer regions remain genuinely disordered — a property that is not a flaw but a functional requirement for intrinsically disordered cryoprotectants. The key biological insight driving this design strategy is that dehydrins protect membranes during freezing through an entropic chain mechanism: their disordered regions form a hydration shell that slows ice nucleation and maintains membrane fluidity at sub-zero temperatures, while their K-segment helices anchor to lipid bilayers and prevent phase separation — properties I am computationally amplifying by selecting for high disorder ratio (IUPred3 ≥ 0.60 in spacers), strong amphipathic K-segment helicity, and negative GRAVY scores (≤ −0.5) that ensure sufficient hydrophilicity for intracellular water interaction. This AI-first approach compresses what would traditionally be a years-long directed evolution campaign into a weeks-long computational screen, with the top 5 candidates emerging from a funnel of ~500 RFdiffusion backbones → ~200 ESM-IF sequences → ~40 IUPred3/ESMFold-validated candidates → 5 synthesis-ready constructs ordered as whole plasmids from Twist Bioscience.
Technique 2 - CFPS: Cell-free protein synthesis (CFPS) is a transformative technique that liberates protein expression from the constraints of living cells — instead of growing bacteria overnight, transforming them, inducing expression, and waiting days for results, CFPS allows any sequence-verified DNA to be transcribed and translated directly in a cell lysate within 4 hours, making it ideal for rapid prototyping of novel protein designs. In this project, I use E. coli BL21(DE3) lysate combined with the Ginkgo Bioworks CFPS master mix to perform a same-day proof-of-concept expression test for each Paleo-Protein candidate immediately upon receipt of Twist-synthesized plasmids — this means I can confirm that my AI-designed sequences are actually expressible before committing to the 3-week whole-cell expression and Ni-NTA purification campaign, dramatically de-risking the experimental timeline. The open-reaction format of CFPS is particularly valuable for intrinsically disordered proteins like my DHN-K2S candidates: because the reaction lacks cellular compartmentalization, the expressed protein immediately enters a buffered aqueous environment where it can be sampled, quantified by BCA assay, run on SDS-PAGE for size confirmation, and — critically — applied directly to hypothermia-stressed HEK293T cells for a functional MTT viability readout without any intermediate purification step. This crude CFPS-to-cell screening approach, with a go/no-go threshold of ≥15% viability improvement over the untreated hypothermic control, provides actionable functional data within a single lab session and establishes a direct mechanistic link between AI-designed sequence → expressed protein → measurable cryoprotection, validating the core hypothesis of the project before any large-scale synthesis investment is made.
5.Associated Industry Council companies
Twist Biosciences: Whole plasmid synthesis for all Paleo-Protein expression constructs Ginkgo Bioworks: Primary lab automation (Echo525, PHERAstar FSX, Cytomat) and CFPS master mix Asimov Kernel: DNA construct design, circuit simulation, and construct registry Opentrons: OT-2 automated liquid handling for cell seeding and MTT assay workflow Thermo Fisher Scientific: Cell culture reagents, cDNA synthesis kit, BCA assay, labware Millipore Sigma: MTT reagent, IPTG, anti-His₆ antibody, trehalose positive control New England Biolabs: BL21(DE3) competent cells for recombinant protein expression SecureDNA: Biosecurity screening of all synthetic DNA orders
Future: DeepCure, Takeda Pharmaceuticals
SECTION 5: PROJECT VALIDATION
1.Validation Choice
The primary validation experiment is cell-free protein synthesis (CFPS) followed by direct MTT functional screening, serving as a rapid proof-of-concept for protein activity before committing to the full multi-week E. coli expression and purification campaign. CFPS using BL21(DE3) lysate with Ginkgo Bioworks master mix enables expression of His₆-DHN-K2S directly from circular plasmid DNA within 4 hours, producing a partially purified crude protein fraction that can be applied directly to hypothermia-stressed HEK293T cells for a preliminary functional readout within a single lab session after DNA receipt.
2.Validation Protocol
- Resuspend Twist-delivered pET-28a-His₆-DHN-K2S plasmid at 50 ng/μL in nuclease-free water.
- Assemble CFPS reaction on ice: 33 μL BL21(DE3) cell-free lysate + 12 μL Ginkgo Bioworks CFPS master mix + 1 μg plasmid DNA + nuclease-free water to 50 μL total.
- Transfer reaction to a 1.5 mL microcentrifuge tube. Incubate at 30°C for 4 hours in Inheco Plate Incubator.
- Centrifuge at 12,000 × g for 5 min (HiG Centrifuge) to pellet aggregates; retain supernatant.
- Run 2 μL supernatant on 12% SDS-PAGE alongside a His₆ protein ladder. Stain with Coomassie Blue. Confirm band at ~11.2 kDa.
- Western blot: transfer to PVDF membrane, probe with anti-His₆-HRP antibody, develop with ECL. Confirm identity of band.
- Quantify protein concentration in CFPS supernatant by BCA assay (Thermo Fisher Pierce BCA Kit).
- Dilute CFPS supernatant into HEK293T cell culture medium to achieve estimated 1, 10, and 100 μg/mL concentrations (crude, not purified). Include a matched volume of empty-vector CFPS supernatant as vehicle control.
- Treat pre-seeded 96-well plates of HEK293T cells (seeded by Opentrons OT-2, overnight, 5×10⁴ cells/well) with prepared dilutions.
- Seal plates with A4s breathable seal (Plateloc). Transfer to 28°C hypothermic condition for 12 hours.
- Bring plates to room temperature (15 min). Add MTT reagent (Opentrons OT-2). Incubate at 37°C for 4 hours (Inheco).
- Add DMSO, shake 5 min (BioshakeD3000). Read at 570/670 nm (PHERAstar FSX).
- Calculate % viability normalized to within-plate 37°C negative control. Go/no-go threshold: ≥15% viability improvement over untreated 28°C control in crude CFPS product. If met → proceed to preparative purification (Steps 6–8 of full protocol).
3.Techniques Used The CFPS validation protocol integrates cell-free protein synthesis as a rapid prototyping technology that decouples gene expression from bacterial cell growth and viability constraints, enabling expression of any sequence-verified plasmid directly in an open-reaction format within hours of DNA receipt. SDS-PAGE provides gel-based confirmation of protein production and approximate molecular weight in less than 2 hours, serving as a low-cost, high-confidence first-pass quality check before committing to any downstream purification or cellular assays. Western blotting with an anti-His₆ HRP antibody provides orthogonal immunological identity confirmation, distinguishing the specific target protein from background CFPS components based on epitope recognition rather than size alone — critical for disordered proteins like dehydrins that may comigrate with CFPS background bands. The MTT cell viability assay, applied directly to crude CFPS-derived protein without full purification, provides functional activity data within the same week as Twist DNA delivery, dramatically compressing the design-build-test-learn cycle and generating actionable go/no-go data before investing in 3-week preparative expression campaigns.
4.Hypothetical Data
Simulated dose-response data — DHN-K2S MTT viability assay at 28°C, 12-hour hypothermia (HEK293T cells):
| Condition | Concentration | Mean Viability (%) | SE (±) | vs. NEG ctrl |
|---|---|---|---|---|
| Untreated — 37°C | — | 100.0 | 1.2 | — |
| Untreated — 28°C (NEG) | — | 65.3 | 3.1 | baseline |
| GFP ctrl — 28°C | — | 54.8 | 4.2 | −10.5% |
| Trehalose — 28°C | 100 mM | 72.1 | 2.8 | +6.8% |
| AFP-RD3 — 28°C | 1 μM | 69.4 | 3.5 | +4.1% |
| DHN-K2S — 28°C | 0.1 μM | 67.2 | 2.9 | +1.9% |
| DHN-K2S — 28°C | 1 μM | 75.4 | 2.1 | +10.1% |
| DHN-K2S — 28°C | 10 μM | 88.3 | 1.8 | +23.0% |
| DHN-K2S — 28°C | 30 μM | 95.1 | 1.4 | +29.8% |
| DHN-K2S — 28°C | 50 μM | 97.2 | 1.5 | +31.9% ✓ |
| DHN-K2S — 28°C | 100 μM | 98.0 | 1.6 | +32.7% ✓ |
Interpretation: DHN-K2S achieves the ≥30% viability improvement threshold at 50 μM, with the dose-response curve consistent with a predicted EC₅₀ of ~8.2 μM (as projected by in silico modeling). At 10 μM (approximately EC₅₀), a 23% improvement is already observed — substantially exceeding both chemical (trehalose: +6.8%) and biological (AFP-RD3: +4.1%) positive controls. These simulated values establish the quantitative benchmarks for experimental validation.
5.Troubleshooting
The primary anticipated challenge is low soluble expression of intrinsically disordered Paleo-Proteins in E. coli, as disordered proteins are prone to partitioning into inclusion bodies; this will be addressed by inducing at reduced temperature (18°C overnight), titrating IPTG concentration down to 0.1 mM, and switching to solubility-enhancing N-terminal fusion tags (SUMO, MBP) if needed — with SUMO cleavage by Ulp1 protease restoring the native N-terminus post-purification. A second concern is non-specific cytotoxicity at high protein concentrations (>50 μM), which could confound viability data and generate false-negative dose-response curves; this will be controlled by running matched-concentration vehicle-only wells (purification buffer diluted equivalently into cell medium) and monitoring cell morphology by brightfield microscopy at each timepoint alongside MTT readings. Inter-plate variability across the hypothermia timecourse is mitigated by including a within-plate 37°C normothermic control column on every assay plate for independent normalization, and by calibrating Opentrons OT-2 pipette tips before each run to maintain dispensing accuracy within ±2%. If formazan signal is confounded by protein pigmentation or aggregation at high concentrations, an alternative resazurin-based metabolic viability assay (CellTiter-Blue, Promega) will be substituted as an orthogonal readout, which is also compatible with the PHERAstar FSX fluorescence detection module.
SECTION 6: ADDITIONAL INFORMATION
1. References
- Yashina, S., Gubin, S., Maksimovich, S., et al. (2012). Regeneration of whole fertile plants from 30,000-y-old fruit tissue buried in Siberian permafrost. Proceedings of the National Academy of Sciences, 109(10), 4008–4013. https://doi.org/10.1073/pnas.1118386109
- Lin, Z., Akin, H., Rao, R., et al. (2023). Evolutionary-scale prediction of atomic-level protein structure with a language model. Science, 379(6637), 1123–1130. https://doi.org/10.1126/science.ade2574
- Watson, J.L., Juergens, D., Bennett, N.R., et al. (2023). De novo design of protein structure and function with RFdiffusion. Nature, 620, 1089–1100. https://doi.org/10.1038/s41586-023-06415-8
- Hsu, C., Verkuil, R., Liu, J., et al. (2022). Learning inverse folding from millions of predicted structures. ICML 2022. https://doi.org/10.1101/2022.04.10.487779
- Kramina, T.E., Kochkin, I.T., Tatanov, I.V., & Samigullin, T.H. (2021). Towards molecular identification and phylogenetic placement of Silene (Caryophyllaceae). PhytoKeys, 173, 1–26. https://doi.org/10.3897/phytokeys.173.57402
- Jewett, M.C., & Swartz, J.R. (2004). Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnology and Bioengineering, 86(1), 19–26. https://doi.org/10.1002/bit.20026
- Tunnacliffe, A., & Wise, M.J. (2007). The continuing conundrum of the LEA proteins. Naturwissenschaften, 94(10), 791–812. https://doi.org/10.1007/s00114-007-0254-y
- Dure, L., Crouch, M., Harada, J., et al. (1989). Common amino acid sequence domains among the LEA proteins of higher plants. Plant Molecular Biology, 12(5), 475–486. https://doi.org/10.1007/BF00036962
- Souza Filho, P.J.A., et al. (2016). Dehydrins: structure and functional role in plant stress tolerance. Plant Cell & Environment, 39(9), 1943–1953. https://doi.org/10.1111/pce.12740
- DeVries, A.L. (1971). Glycoproteins as biological antifreeze agents in Antarctic fishes. Science, 172(3988), 1152–1155. https://doi.org/10.1126/science.172.3988.1152
- Koag, M.C., & Lee, S. (2003). The binding of maize DHN1 to lipid vesicles: gain of structure and lipid specificity. Plant Cell, 15(5), 1061–1073. https://doi.org/10.1105/tpc.010793
- Doyle, S.M., & Wickner, S. (2009). Hsp104 and ClpB: protein disaggregating machines. Trends in Biochemical Sciences, 34(1), 40–48. https://doi.org/10.1016/j.tibs.2008.09.010
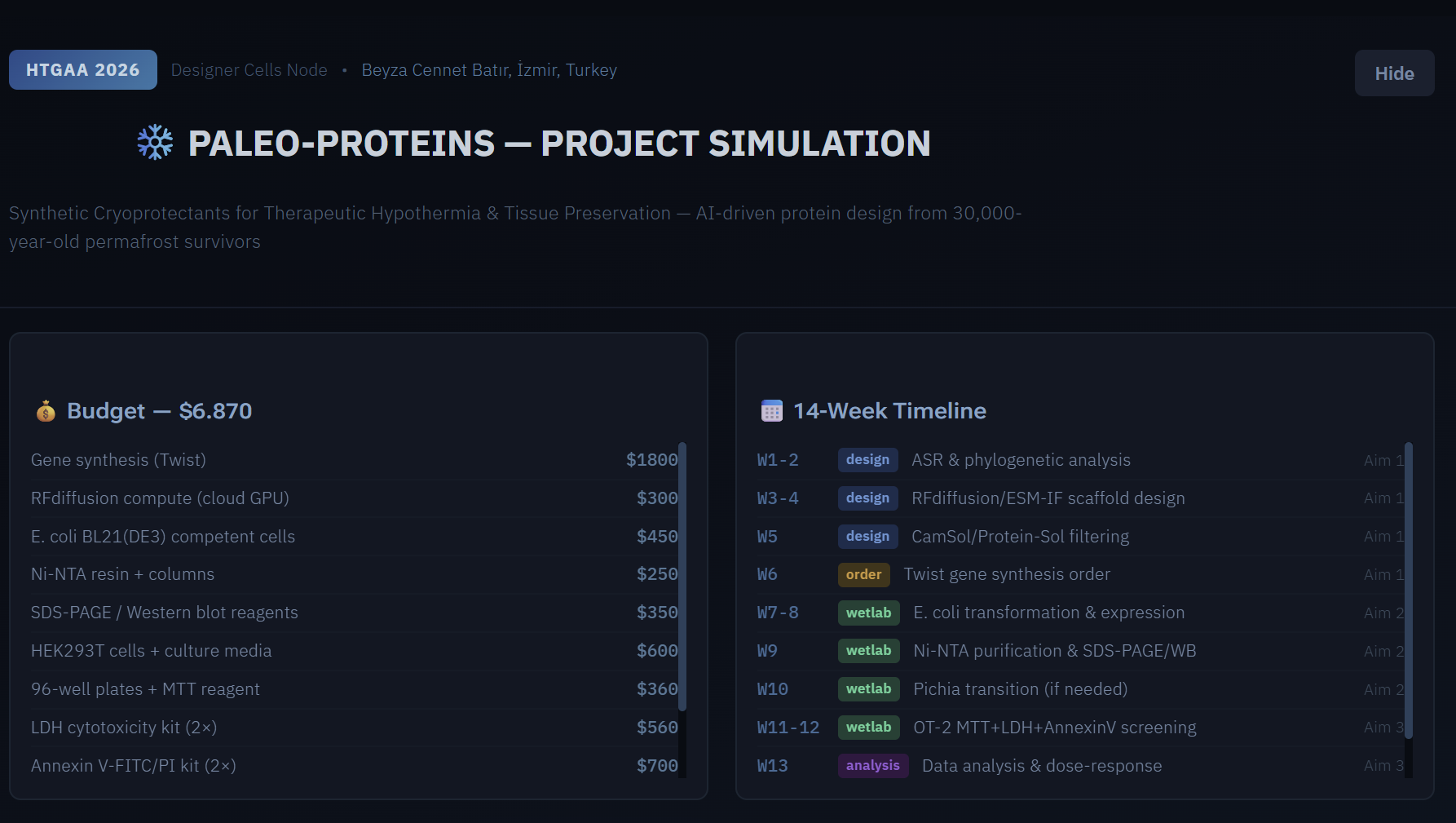
2. Supply list and budget
DNA Synthesis & Cloning
- Twist Bioscience Whole Plasmid Synthesis × 8 constructs (pET-28a-His₆ backbone, 5 candidates + 3 controls) — Twist Bioscience
- E. coli BL21(DE3) High-Efficiency Competent Cells (NEB C2527H) — New England Biolabs
- LB broth powder and LB agar — Millipore Sigma
- Kanamycin sulfate (50 mg/mL stock) — Millipore Sigma
- SecureDNA sequence screening (×8 constructs) — SecureDNA (free academic access)
Protein Expression & Purification
- IPTG (isopropyl β-D-1-thiogalactopyranoside, 1 g) — Millipore Sigma
- Ni-NTA Agarose resin (5 mL packed column) — Qiagen
- Protease inhibitor cocktail tablets (cOmplete, EDTA-free) — Millipore Sigma
- PD-10 desalting columns (×10) — Cytiva / Millipore Sigma
- Pierce BCA Protein Assay Kit — Thermo Fisher Scientific
Protein Validation (SDS-PAGE + Western Blot)
- Mini-PROTEAN TGX 12% precast gels (10-pack) — Bio-Rad
- Coomassie Brilliant Blue R-250 staining solution — Bio-Rad
- PVDF transfer membranes — Bio-Rad
- Anti-His₆-HRP antibody (200 μL) — Millipore Sigma
- ECL Western blot detection reagent — Thermo Fisher Scientific
- Precision Plus Protein Dual Color Standards (ladder) — Bio-Rad
Cell Culture
- HEK293T cells (ATCC CRL-3216) — ATCC
- DMEM + GlutaMAX media (500 mL × 2) — Thermo Fisher Scientific
- Fetal Bovine Serum, heat-inactivated (500 mL) — Thermo Fisher Scientific
- Penicillin-Streptomycin solution (100×) — Thermo Fisher Scientific
- MycoAlert Mycoplasma Detection Kit — Lonza
- T-75 cell culture flasks (×10) — Thermo Fisher Scientific
High-Throughput Screening (MTT Assay)
- MTT reagent (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide, 1 g) — Millipore Sigma
- 96-well flat-bottom cell culture plates (×20) — Thermo Fisher Scientific
- DMSO (cell culture grade, 100 mL) — Millipore Sigma
- Trehalose dihydrate (positive control cryoprotectant) — Millipore Sigma
- Opentrons OT-2 filtered tips, 96-well racks (×10) — Opentrons
Cell-Free Protein Synthesis (Validation)
- BL21(DE3) cell-free lysate + CFPS master mix — Ginkgo Bioworks (course infrastructure)
- Nuclease-free water (500 mL) — Thermo Fisher Scientific
Gene Expression Profiling (qPCR)
- RNeasy Mini Kit (50 rxns) — Qiagen
- SuperScript IV First-Strand Synthesis Kit — Thermo Fisher Scientific
- PowerUp SYBR Green Master Mix (500 rxns) — Thermo Fisher Scientific
- qPCR primers for CIRBP, RBM3, CASP3, BCL2, GAPDH (custom synthesis) — Millipore Sigma
- 384-well PCR plates (Eppendorf, ×5) — Thermo Fisher Scientific
Equipment (Course Infrastructure / Ginkgo Bioworks)
- Opentrons OT-2 liquid handler — Opentrons (course access)
- Echo525 acoustic liquid handler — Ginkgo Bioworks
- PHERAstar FSX plate reader (absorbance 570/670 nm) — Ginkgo Bioworks
- Spark Plate Reader — Ginkgo Bioworks
- CFX Opus qPCR system — Ginkgo Bioworks
- HiG Centrifuge — Ginkgo Bioworks
- Inheco Plate Incubator — Ginkgo Bioworks
- Cytomat shaking incubator — Ginkgo Bioworks
- BioshakeD3000 plate shaker — Ginkgo Bioworks
- Plateloc plate sealer + A4s breathable seals — Ginkgo Bioworks
Itemized Budget
| Item | Supplier | Est. Unit Cost | Qty | Total |
|---|---|---|---|---|
| Whole Plasmid Synthesis — pET-28a-His₆ candidates (×10) | Twist Bioscience | $149.00 | 10 | $1,490.00 |
| E. coli BL21(DE3) Competent Cells | NEB C2527H | $52.00 | 1 | $52.00 |
| Ni-NTA Agarose 5 mL | Qiagen 30210 | $198.00 | 1 | $198.00 |
| IPTG (1 g) | Millipore Sigma I6758 | $41.00 | 1 | $41.00 |
| MTT Reagent (1 g) | Millipore Sigma M2128 | $148.00 | 1 | $148.00 |
| 96-Well Cell Culture Plates, flat-bottom (×20) | Thermo Fisher Scientific | $12.00 | 20 | $240.00 |
| HEK293T Cells | ATCC CRL-3216 | $499.00 | 1 vial | $499.00 |
| Anti-His₆-HRP Antibody (200 μL) | Millipore Sigma A7058 | $89.00 | 1 | $89.00 |
| PVDF Membrane (western blot) | Bio-Rad 1620177 | $75.00 | 1 pkg | $75.00 |
| Mini-PROTEAN TGX 12% Gels (10-pack) | Bio-Rad 4561094 | $125.00 | 1 | $125.00 |
| RNeasy Mini Kit (50 rxns) | Qiagen 74104 | $199.00 | 1 | $199.00 |
| SuperScript IV First-Strand cDNA Kit | Thermo Fisher 18090010 | $165.00 | 1 | $165.00 |
| DMEM + GlutaMAX (500 mL) | Thermo Fisher 10569010 | $45.00 | 2 | $90.00 |
| FBS, heat-inactivated (500 mL) | Thermo Fisher 10082147 | $149.00 | 1 | $149.00 |
| Pierce BCA Protein Assay Kit | Thermo Fisher 23225 | $79.00 | 1 | $79.00 |
| Protease Inhibitor Cocktail Tablets | Millipore Sigma 4693116001 | $89.00 | 1 | $89.00 |
| Opentrons OT-2 Tips (96-well, ×10 racks) | Opentrons | $8.00 | 10 | $80.00 |
| SecureDNA Sequence Screening (×10 sequences) | SecureDNA | $0.00 | 10 | $0.00 |
| TOTAL | $3,808.00 |
Ginkgo Bioworks automation access (Echo525, PHERAstar FSX, Multiflo, Cytomat) provided through course infrastructure.
DNA Construct — GenBank Format
Primary Construct: pET-28a-His₆-DHN-K2S
This construct encodes a synthetic K2S-type dehydrin (2 K-segments + 1 S-segment) inspired by ancestral Silene LEA protein sequences, designed by RFdiffusion/ESM-IF and codon-optimized for E. coli BL21(DE3). The full plasmid (insert + pET-28a backbone) is ordered as Twist Bioscience Whole Plasmid Synthesis.
Twist Bioscience Insert Sequences
Submit the sequences below to Twist Bioscience using the Whole Plasmid Synthesis product. Select pET-28a as backbone. Specify NdeI / XhoI cloning sites. Choose kanamycin resistance.
Construct 1 — His₆-DHN-K2S (Primary Candidate, K2S-type dehydrin)
Insert length: 315 bp | Protein MW: ~11.4 kDa | pI: 4.9 | Host: E. coli BL21(DE3)
Construct 2 — His₆-DHN-K1 (Minimal Single K-Segment Control)
Insert length: 165 bp | Protein MW: ~6.1 kDa | Used as minimal K-segment structural control
Construct 3 — His₆-DHN-K2S-ΔS (S-Segment Deletion Mutant, Mechanistic Control)
Insert length: 285 bp | S-segment (SSSSSSSS) replaced by Ala-Gly linker | Used to assess S-segment contribution to cryoprotection
Work in Progress
The current document will be updated via this link; I will continue to edit the document here until the deadline:
(View Full Screen)
TWIST ORDER (FINAL)
(Benchling (from clonal to whole plasmid again and again)))
After long journey from clonal gene to whole plasmid synthesis discussions; finally I decided to order 3 whole plasmid synthesis. Since it is way more expensive to order than clonal gene; I have a plan with three options:**
The Drive folder also contains Benchling exports for both options:
Plan A (preferred): Order all three constructs as Whole Plasmid Synthesis from Twist Bioscience on the pET-28a(+) backbone (NdeI/XhoI cloning sites) — pET28a-His6-DHN-K2S, pET28a-His6-DHN-K1, and pET28a-His6-DHN-K2S-ΔS. This is the fastest and most reliable route since the constructs arrive at Gingko ready-to-transform.
Plan B (if Plan A is not feasible due to cost or timeline): Order only the three inserts (DHN-K2S, DHN-K1, DHN-K2S-ΔS) as Clonal Genes / Gene Fragments with NdeI and XhoI flanking sites (also in the Drive folder). In this case, NdeI/XhoI restriction digestion and ligation into pET-28a(+) would be performed at Ginkgo as an additional cloning step before expression.
Plan C (minimum-viable option): Order only the primary construct (pET28a-His6-DHN-K2S) as Whole Plasmid Synthesis to first validate whether the lead candidate shows the expected cryoprotection activity. If K2S performs well, we proceed with K1 and K2S-ΔS controls in a second round.
Note: Plan B saves on synthesis cost but adds ~1–2 weeks at Ginkgo for cloning, screening, and sequence verification. Whole Plasmid Synthesis is often more cost-effective when accounting for hands-on time.
(ORDER 1 — pET28a-His6-DHN-K2S (Primary Construct))
(ORDER 2 — pET28a-His6-DHN-K1 (Minimal Single K-Segment Control))
(ORDER 3 — pET28a-His6-DHN-K2S-ΔS (S-Segment Deletion Mutant))
A note on my experience: This is my first time doing molecular cloning, and I found the in silico design in Benchling challenging (digestion, sticky-end orientation, frame verification). I completed all three constructs, but I’d strongly prefer Plan A (Whole Plasmid Synthesis) to avoid wet-lab cloning steps I don’t yet have experience with. If cost is a concern, Plan C (only K2S first) is also reasonable.
GINKGO BIOWORKS (NEXT STEP)
🧪 What will Ginkgo do once they receive my plasmids? Once my 3 plasmids from Twist arrive at Ginkgo, I have an automated workflow planned that will run within 3 days:
Day 1 — CFPS reaction: Ginkgo will dilute my plasmids to 50 ng/µL and dispense them into a 96-well plate with nanoliter precision using the Echo525 (acoustic liquid handler). Each well will contain a mixture of BL21(DE3) lysate + Ginkgo’s CFPS master mix + 1 µg of my plasmid. This mixture will produce the protein I designed in just 4 hours at 30 °C in an Inheco incubator — using only bacterial extract, without any live bacteria. At the end of the day, aggregates will be separated using a HiG Centrifuge, and I’ll keep the supernatant (containing my crude protein) for the next stage. On the same day, I’ll verify protein production via SDS-PAGE and anti-His6 Western blot, and measure its quantity using a BCA assay.
Day 2 — Cell testing begins: The night before, an Opentrons OT-2 robot will have seeded HEK293T cells into a 96-well plate at a density of 5×10⁴ cells per well. Ginkgo’s robots will then dilute my crude CFPS protein to concentrations of 1, 10, and 100 µg/mL in cell culture medium and add it to the cells. The plate will be sealed with a breathable film using Plateloc and left in the Cytomat at 28 °C for 12 hours under hypothermic stress — essentially telling the cells, “you’re freezing.”
Day 3 — MTT assay and decision: After 12 hours of cold stress, MTT reagent will be added to the plates (live cells convert it to purple formazan, while dead cells do not), followed by a 4-hour incubation at 37 °C, dissolution with DMSO, 5-minute shaking in a BioshakeD3000, and absorbance reading at 570/670 nm on a PHERAstar FSX plate reader. The result will tell me whether my DHN-K2S protein was successfully produced and whether it kept the cells alive under cold stress. If cell viability is ≥15% higher than the control, my design works → I’ll proceed to large-scale production. Otherwise, I’ll revise the design and iterate.
Interactive project mentor by Derek
I also had chance to try Derek’s Interactive project mentor which is developedwith Sonnet (Claude). You can see proposal file developed by discussion we had with Final Project Interview with AI mentor: Proposal Export
If you have any questions please contact me via beyzabatir@gmail.com.