Individual Final Project
iKe
Glacier Preservation


Abstract
Mountain glaciers are melting progressively due to climate change and human activity. I am inspired by the glaciers of the Italian Alps where my family is from and throughout generations have seen firsthand the glaciers progressively disappear. Glaciers are vital ecosystems which contribute to protecting nature and human existence.
I aim to use my knowledge in textiles, biology and biodesign to help preserve and rebuild glaciers using ice nucleating proteins (INPs).
I theorise that inoculating glaciers with modified INPs using cell free synthesis would help improve ice catalyzation and make glacier ice more resilient to face increasing temperatures. This method will work as a defensive tool helping restore natural and healthy glacier cycles benefiting a wider ecosystem and battling climate change.
Project overview
Growing up I spent a lot of time in the Italian Alps where part of my family originates from, within only 23 years of life I have seen first hand the glaciers of mountains surrounding me disappear progressively, my mother sees an even bigger decrease and my grandfather a shocking decrease. With my background in textiles, my current studies in Biodesign, my curiosity for biology and now partaking in HTGAAA I will conceptualise a project combining textiles and biology as a means to create a tool which could help prevent the fast disappearance of glaciers.
Glaciers are vital elements to regulating the earth’s temperature, they are ecosystems of their own across the world. As pointed out by Glacier Preservation, “glaciers are essential for sustaining millions of people by providing fresh water, supporting hydropower generation, and playing a key role in environmental stability. However, as climate change accelerates, glaciers are retreating at an unprecedented rate, threatening water security, energy infrastructure, and increasing natural hazards like flooding and avalanches.” (Un-glaciers.org. (2026). Glacier Preservation is the Key to Ensuring the Security of Water, Energy, and Environmental Resources. [online] Available at: https://www.un-glaciers.org/en/articles/glacier-preservation-key-ensuring-security-water-energy-and-environmental-resources.).
Glaciers are rapidly melting and disappearing but it is continuously overlooked, as explained by the Geneva Environment Network “Ice mass covers 10% of the Earth land surface (Antarctic ice sheet 8.3%, Greenland ice sheet 1.2%, glaciers and ice caps 0.5%), and its loss is a primary marker of climate change. The decrease has accelerated in recent decades and is now reaching concerning levels”. This issue will lead to (and has already started) ecosystem health, biodiversity health, human health and social cultural repercussions as well as a global increase of climate change consequences. Mountain glaciers are first most affected by glaciers disappearance, Alpine glaciers are expected to lose 90% of their mass by 2100 according to Harry Zekollari, 2019 and a third of global glaciers of World Heritage are predicted to have disappeared by 2050 according to the UNESCO in 2022, we are in 2026. (Please find below more complementary information)
My project aims to ethically preserve glaciers and help them naturally rebuild while respecting their natural cycles and having minimal interference. I want to work with ice nucleating proteins, specifically the inaK strands, they are commonly found in nature and have the function to catalyze ice. These INPs should be used to improve ice formation in glaciers and sustain ice levels while they face a rise in temperatures. I aim to work using a cell free synthesis method, potentially improve the inaK function through gene mutation and through different levels of test innoculate glaciers with inaK.
The following graphs and analysis by Geneva Environment Network depict the intense loss of mass of glaciers.
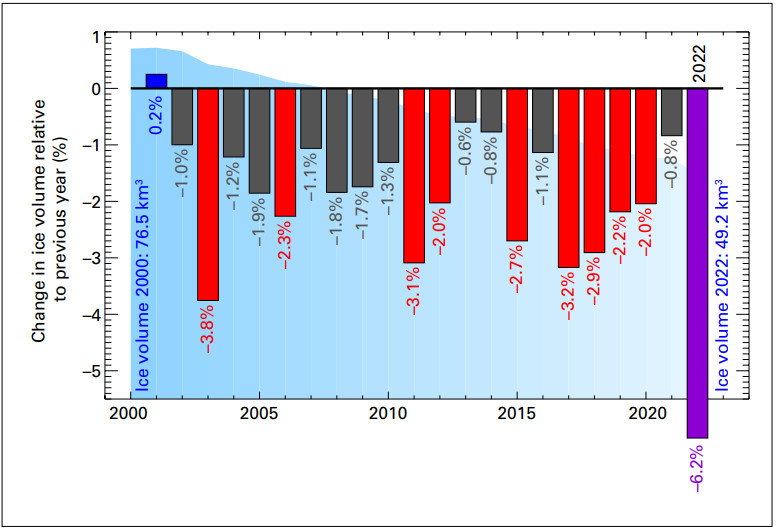
“Total annual loss 2022 of Swiss glaciers related to the current ice volume 2002–2022. The vertical bars indicate the percentage change in ice volume relative to the previous year. Red and purple bars are the 10 largest relative mass losses on record. The purple bar is the relative mass loss for 2022. The blue-shaded area in the background represents the overall ice volume. Source: Matthias Huss based on Glacier Monitoring Switzerland, 2022: Swiss Glacier Mass Balance (2022). The summer of 2023 was the second-most negative year in history of Swiss glaciers retreat, with glaciers clearly below the average of the last 10 years. With 4% of ice volume destroyed in 2023, the Swiss Commission for Cryosphere Observation of the Swiss Academy of Sciences reported that a total of 10% of the ice volume disappeared in only two years.”
“Glaciers in Switzerland lost half their volume between 1931 and 2016 and another 12% between 2016 and 2021. This ice loss was described in a study published in the scientific journal The Cryosphere in August 2022 by a team of researchers from ETH Zurich and the Swiss Federal Institute for Forest, Snow and Landscape Research (WSL). The World Meteorological Organization 2022 State of Climate Report, signals that between 2021 and 2022, an additional 6% of glacier ice volume was lost.” (“Unprecedented Rates of Mountain Glacier Melting | Glaciers and the Role of Geneva”)
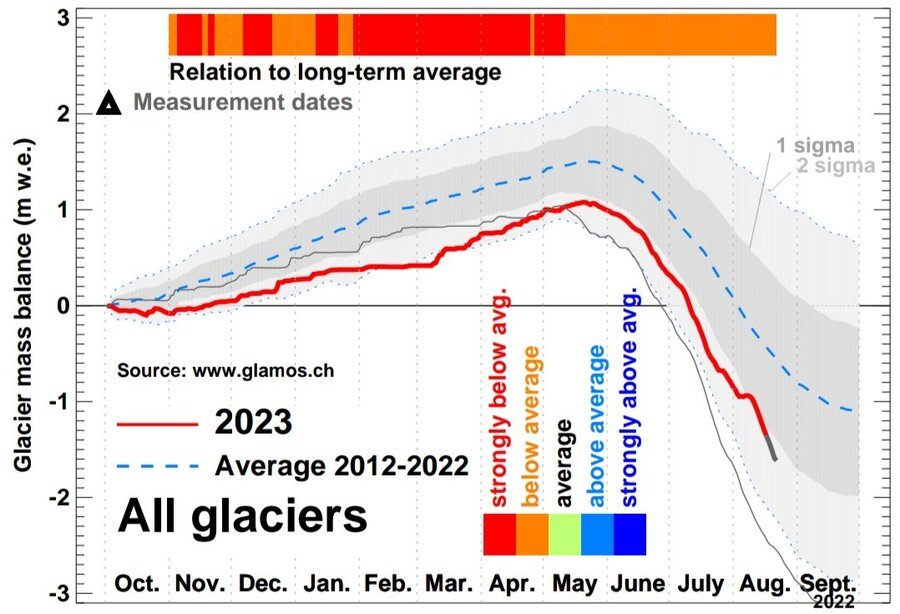
Source: GLAMOS – Glacier Monitoring in Switzerland
“Measurations undertaken by GLAMOS at the end of the winter season of 2024 found strongly above-average snow cover on glaciers in all regions of Switzerland with average snow depths of 3 to 6 meters. Extrapolated to all Swiss glaciers, a surplus of 31% more winter snow compared to the period 2010-2020 is found. After the very dry winters 2022 and 2023, with corresponding extreme ice loss in summer, the abundant snow falls during winter 2024 represent a blessing for glaciers. Despite exceptionally large volumes of snow last winter, during the summer of 2024, Swiss glaciers have lost 2.5% of their volume, with data recorded by the Glacier Monitoring in Switzerland (GLAMOS) network finding that the retreat of the glacier tongues and their disintegration, a clear symptom of climate change, has caused in 2022 and 2023 a total of 10% of Swiss glacier volume to disappear, with losses in 2024 exceeding the mean value of the last decade.” (“Unprecedented Rates of Mountain Glacier Melting | Glaciers and the Role of Geneva”)

“Unprecedented Rates of Mountain Glacier Melting | Glaciers and the Role of Geneva.” Genevaenvironmentnetwork.org, 2024, www.genevaenvironmentnetwork.org/fr/ressources/nouvelles/unprecedented-rates-of-mountain-glacier-melting/.
Project Aims
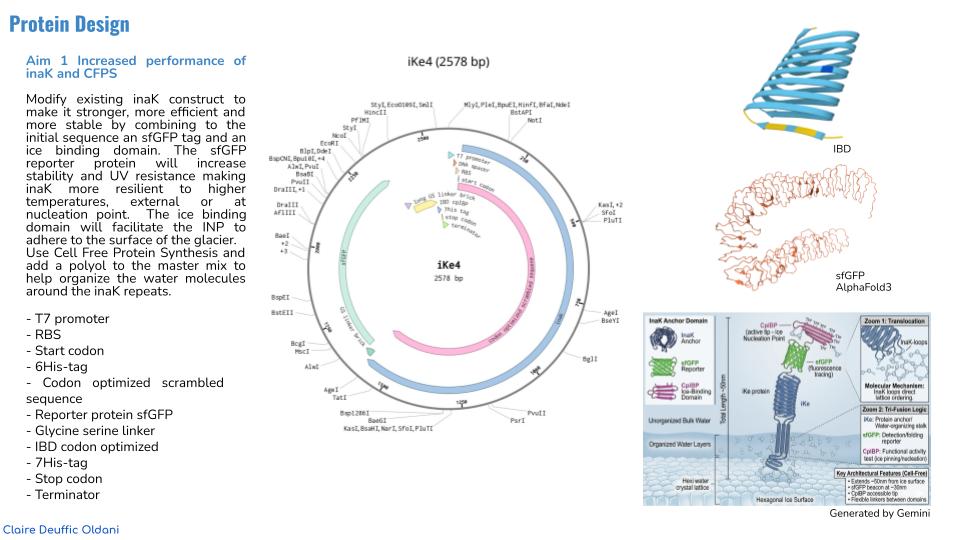
Aim 1 Increased performance of inaK and CFPS
Modify existing inaK construct to make it stronger, more efficient and more stable by combining to the initial sequence an sfGFP tag and an ice binding domain. The sfGFP reporter protein will increase stability and UV resistance making inaK more resilient to higher temperatures, external or at nucleation point. The ice binding domain will facilitate the INP to adhere to the surface of the glacier. Use Cell Free Protein Synthesis and add a polyol to the master mix to help organize the water molecules around the inaK repeats.
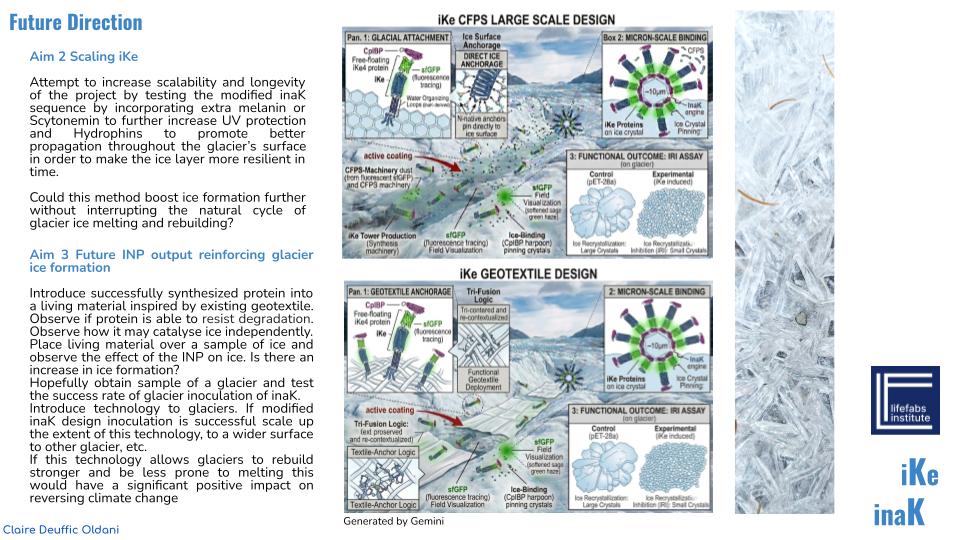
Aim 2 Scaling inaK
Attempt to increase scalability and longevity of the project by testing the modified inaK sequence by incorporating extra melanin or Scytonemin to further increase UV protection and Hydrophins to promote better propagation throughout the glacier’s surface in order to make the ice layer more resilient in time.
Could this method boost ice formation further without interrupting the natural cycle of glacier ice melting and rebuilding?
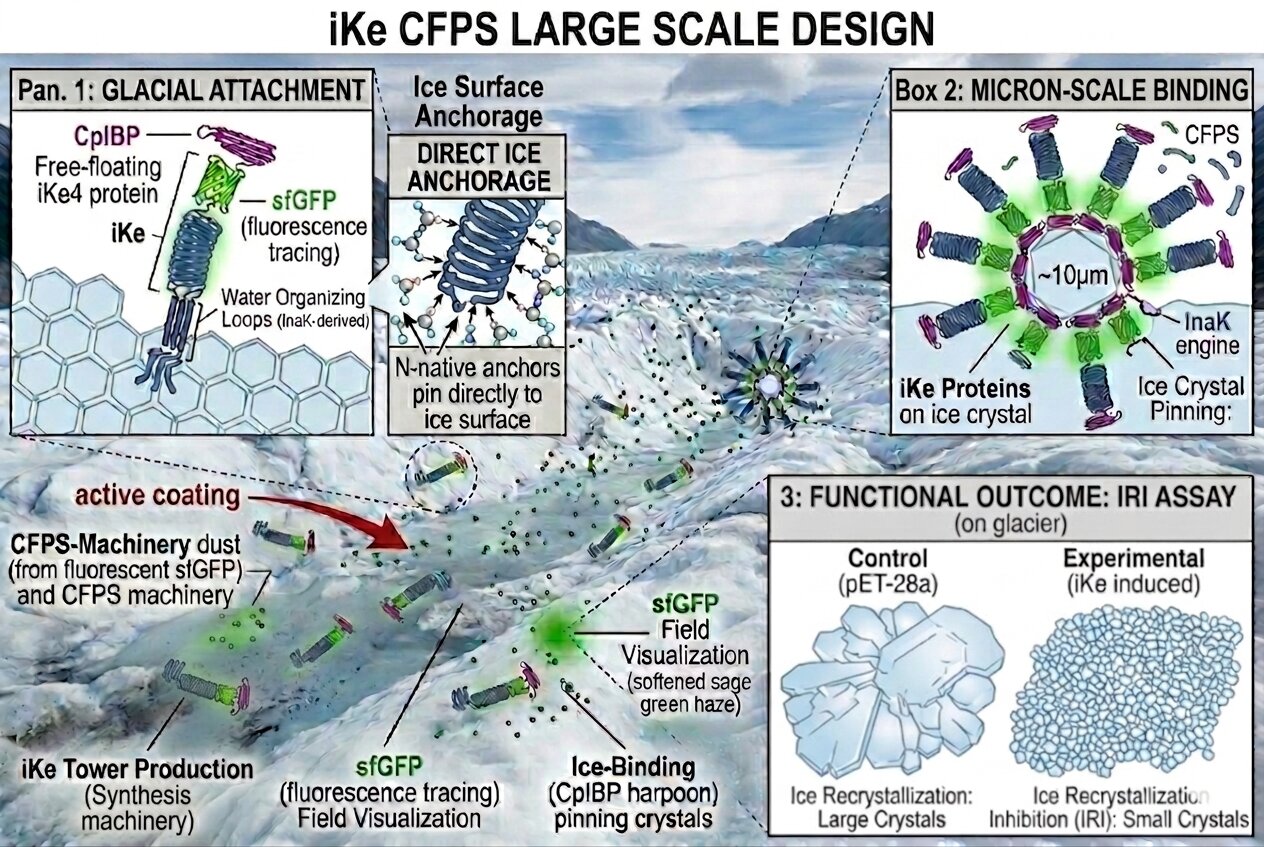
Methods to apply iKe technology to glaciers at this point of research
Similarly to cloud seeding one could disperse the iKe technology using a bio-aerosol spraying technique or a pellet method. These applications would allow to cover a large surface of the glaciers efficiently with low impact.
Aim 3 Future INP output reinforcing glacier ice formation
Introduce successfully synthesized protein into a living material inspired by existing geotextile. Observe if protein is able to resist degredation. Observe how it may catalyse ice independently. Place living material over a sample of ice and observe the effect of the INP on ice. Is there an increase in ice formation? Hopefully obtain sample of a glacier and test the success rate of glacier inoculation of inaK. Introduce technology to glaciers. If modified inaK design inoculation is successful scale up the extent of this technology, to a wider surface to other glacier, etc. If this technology allows glaciers to rebuild stronger and be less prone to melting this would have a significant positive impact on reversing climate change
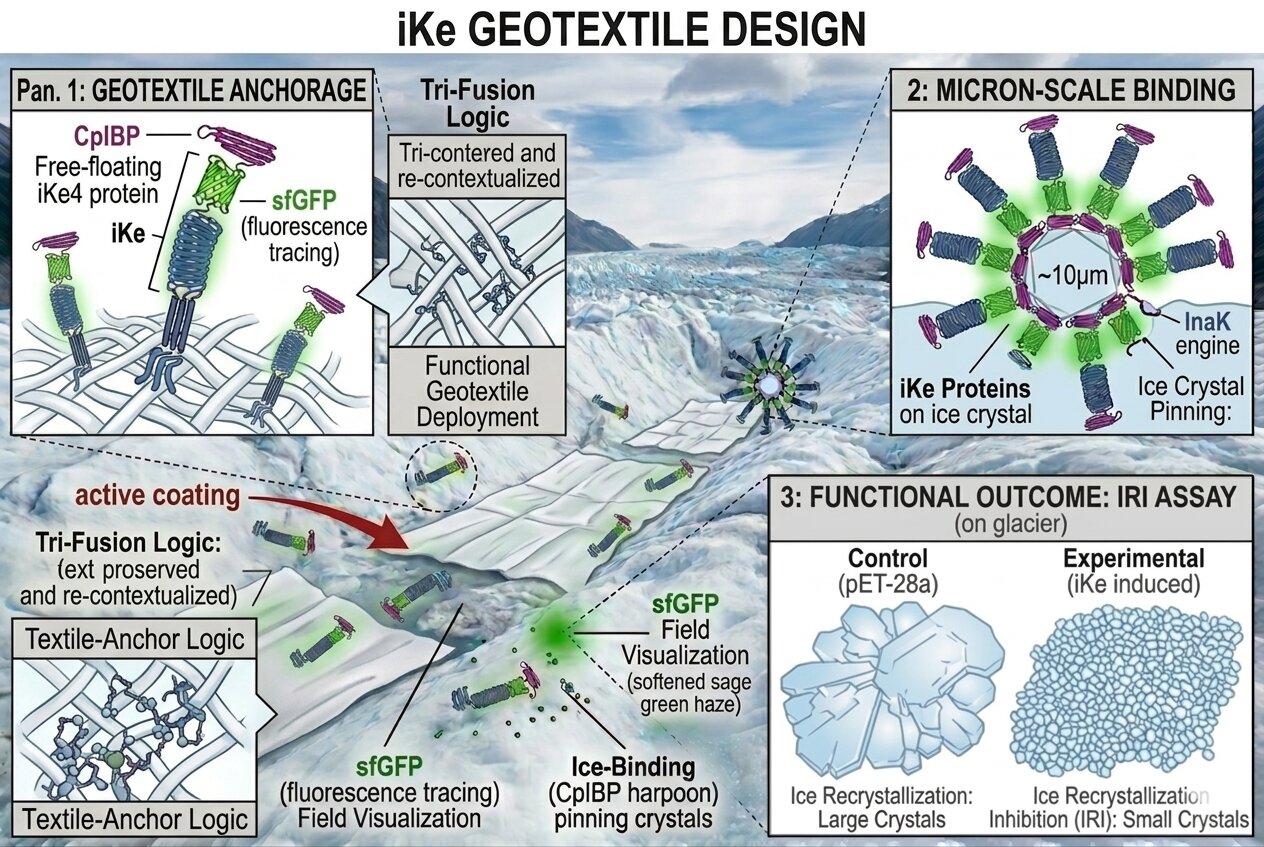
Future direction
If I were to select a material to use for my aim three I would choose mycelium. I could work with “deactivated” mycelium which has been pressed with heat and stunts its growth offering better control of the material, if working with living mycelium it would require nutrition but mostly would simply enter a dormant state in freezing conditions. Mycelium is a wonderful alternative to regular synthetic geotextiles, it would limit impact on local ecosystems and is biodegradable if needed. Mycelium also offers excellent insulation as it forms many air pockets which could further help protect ice mass of glaciers and the new provided ice crystallization as they face warm temperatures. On one hand, I could inoculate my material fully with my protein, the mycelium is a beneficial material to ice nucleation as it is very porous, allowing water to enter deep in its layers, reaching all ice nucleating proteins and allowing them to execute their function. On the other hand, I could inoculate my ike technology using a surface loading method like a textile anchor logic working with material affinity binding, in this case it could involve adding a cellulose binding domain for the proteins to “stick” to my material. Additionally, mycelium could potentially provide a strong scaffold for ice crystallization as it is a very fibrous organism, offering many anchoring points for aggregation and ice lattice increasing the density of ice formation compared to regular flat surfaced synthetic fibers.
Note that if CFPS fails, then attempt in vivo synthesis and introduce inaK to new host, Pseudomonas Fluorescens, which does not contain any ice-nucleotides, already exist within glaciers and presents no risk to nature. Transcribing new DNA data from one Pseudomonas to another will be either as they are part of the same species. Additionally, because Pseudomonas Fluorescens already exist within glaciers it facilitates horizontal gene transfer but also avoids disrupting the natural balance and ecosystem of the glaciers.
Within this project I will also focus a section of my research in ethics, analyzing the impact my work could have on glaciers and their ecosystems. I aim to be educated about environmental laws and bioethics.
I recognise that within the context of this project I would need the expert knowledge of a glaciologue able to provide me with specific and reliable data to use within my research which would allow me to work as ethically, sustainably and durably as possible.
Background & Literature Context
Peer Review Research Citations
“Ice-nucleation active (INA) bacteria can promote the growth of ice more effectively than any other known material. Using specialized ice-nucleating proteins (INPs), they obtain nutrients from plants by inducing frost damage and, when airborne in the atmosphere, they drive ice nucleation within clouds, which may affect global precipitation patterns” Roeters, S.J., Golbek, T.W., Bregnhøj, M., et al. (2021) ‘Ice-nucleating proteins are activated by low temperatures to control the structure of interfacial water’, Nature Communications
The opening line from Roeters in ‘Ice-nucleating proteins are activated by low temperatures to control the structure of interfacial water’ (2021), briefly shows the potential of ice nucleating proteins, it depicts very clearly that ice nucleating proteins (INPs) can “create” ice, or rather increase the catalyzation of ice. These INPs are commonly found in bacterias such as Pseudomona Synrigae which is classified as pathogenic as it contains INPs providing it with the capacity to freeze crops. INPs can act on a small scale in ice nucleating active bacteria as well as on a larger scale (as I will explore further in throughout my project) demonstrating potential of application as INPs affect natural systems on a whole. Overall, this citation and article explains how INPs increase the production of ice “more effectively than any other known material” to science as they have a unique way of organizing water molecules.
“Modeling of Pseudomonas borealis INP by AlphaFold suggests that the central domain of 65 tandem sixteen-residue repeats forms a beta-solenoid with arrays of outwardpointing threonines and tyrosines, which may organize water molecules into an ice-like pattern.” Forbes, J., Bissoyi, A., Eickhoff, L., et al. (2022) ‘Water-organizing motif continuity is critical for potent ice nucleation protein activity’, Nature Communications
This citation and study from Forbes explains that the long continuous and repetitive nature of an INP domain is key to effective ice nucleation, it provides a surface to organize water molecules and form ice lattice. Furthermore, it demonstrates that disrupting the continuous motif by modifying the sequence, adding a construct with the sequence or changing the size of the sequence can lead to reducing nucleation function.
Literature review
There are a few ongoing projects which aim to protect glaciers. Mainly they appear in the form of geotextile covering. GlacierProtect by Naue is a glacier protection project through geotextiles, it uses sustainable raw materials to reflect up to 75% of sunlight. Glaciers already have the capacity to reflect sunlight but this new textile supports this natural ability which preserves the glaciers and by preserving glaciers will enable them better form and themselves better reflect the sunlight. This project inspires me as a way of demonstrating these innovative projects are scalable but also that true regenerative design can be achieved with a mindset of supporting glaciers to promote their rebuild and growth where in turn they could thrive and function as they were meant to. Ponte di Legno Tonale glacier project with a similar technology to Naue, the Presena glacier has been protected since 2008 after observing significant damage to the glacier in 2000 by “covering Presena glacier with geotextile fabric covers, which are able to reduce by 50% the melting of snow and ice during summer months” (Ponte di Legno Tonale, n.d.). This project demonstrating chronological progress shows the real potential and benefit of protecting glaciers and assisting them to rebuild naturally. Furthermore, many companies are innovating and making use of ice nucleating proteins in snow canons to increase production of snow. Snowmax International has made great progress in this direction by using INP to make water free at higher temperatures because of a higher concentration of nuclides, increase ice catalyzation speed as it attaches to nuclides, and be more resistant in time as it makes use of less water therefore reducing evaporation capacity and due to its smaller crystal structures from stronger iconic bonds. According to GoldBio, “P. syringae’s ice-nucleating activities have long been used to make artificial snow. Products including Snomax® use the proteins derived from the outside of bacteria to enhance the snow generated by snow blowers. One study showed that Snomax® increases the amount of snow made by a snow blower by as much as 90% (Snomax® International, 2013)” (Christner et al., 2008) leading to my next point of research. While this is mostly destined to ski track upkeep and not oriented to ice production it demonstrates that INPs are being used on a larger scale and prove to be effective. These technologies are also proving that they have no negative repercussions on the environment as they make use of all natural compounds. Pseudomonas syringae is a bacteria which contains ice nucleating protein strands which have the ability to catalyze the formation of ice in sub zero temperatures. The main issue with pseudomonas syringae is that it is a pathogen in agriculture as it infects crops. However, would it be possible to extract the ice nucleating protein strands from the bacteria, use pseudomonas syringae as a model to synthesize ice nucleating protein or genetically modify the bacteria to neutralize its harmful effect to plants in order to use this genetically modified or synthesised bacteria as an ice forming agent to help preserve glaciers. Instant ice packs are innovations of modern medicine where a sealed pouch of water is activated through an endothermic chemical reaction transforming surrounding heat into ice. Would it be possible to upscale this chemical reaction or let it inspire some biomimicry which through textiles and biology could use the increasingly hot temperature of the earth to transform it into ice on the surface of glaciers to help preserve them. Snow Seeds by Tanay Wadokar is a project presented by a 2025 graduate of the MA Materials Future at Central Saint Martins (UAL, London). Tanay Wadokar created snow board stickers with cloud seeding technology. This would allow the deposit of ice nuclei while snowboarding which would enable locals and tourists to enjoy the inter sport while preserving mountains and glaciers by regenerating snowfalls. It is not about limiting the life we know today but rather by shaping it in a sustainable proactive way.
Ice nucleation is a mechanism where water molecules transform from a liquid state to a solid crystalline lattice, an existing nucleus or template needs to surpass the energy barrier of crystallization.
In a pure water sample, containing no foreign particles, water molecules can spontaneously collide and form stable crystals, this is homogeneous nucleation. In contrast a heterogeneous nucleation is when a foreign compound, an ice nucleator, forms a physical surface or template for the water molecules to attach themselves to which significantly lowers the energy needed for freezing. In the case of Pseudomonas syringae the INPs provide a heterogeneous surface for ice to form itself on. Ice nucleation temperature will depend on the context in which ice nucleation is occurring, typically for pure water samples can stay liquid till -40°C in a supercooled state before freezing. With common nucleators such as minerals the water sample can freeze at about -10°C to -15°C. In the case of the inaK INP which I am studying, the ice catalyzation can be triggered at -2°C to -5°C, demonstrating the potential of using inaK compared to other INP. Being able to catalyze ice at higher subzero temperatures shows potential for environmental protection, as seen in the Snowmax project, helping to produce snow at higher temperatures, essential for protecting glaciers and reducing the ice loss. Additionally, they can have an atmospheric impact, because INPs are so effective they can truly affect weather patterns, similarly to cloud seeding technology they can trigger cloud formation and influence precipitation at warmer altitudes. For companies such as Snowmax the ice nucleation technologies allow them to produce the same amount of snow using less energy than they would need at regular ice nucleation temperature of -10°C or -15°C as machines don’t need to compensate for the temperature.
Different types of ice nucleation particles exist, they can be organized in categories.
The mineral particles which are mostly inorganic atmospheric aerosols like minerals and soot. They are the most common INPs in the atmosphere but are not considered very efficient as they require the very low temperatures of -10°C to -15°C to initiate heterogeneous nucleation. The biological ice nucleators which consist of bacteria such as the Pseudomonas syringae containing inaK. They are the most effective ice nucleators as they can trigger ice at higher temperatures, between -2°C and -5°C. Like the inaK protein they are usually very efficient due to their repetitive genetic code improving crystal lattice capability. Organic and macromolecular compounds which refer to non-living organic substances. Polyols are an example of macromolecular compounds which are not always ice nucleators but can improve the efficiency of existing biological nucleators by up to 100 fold. Engineered materials, however, are synthetic substances designed to imitate natural ice nucleation. Silver iodide for instance is a synthetic protein used in cloud seeding. Biological nucleators such as inaK are the most relevant for my work as, on one hand, they prove to be more efficient for ice nucleation at higher sub zero temperatures which will be essential when working on glaciers confronted with rising temperatures. On the other hand, they have a better chance of non disruptive inoculation into glaciers as they are more likely to assimilate naturally to the existing biological nucleators already present in glaciers.
Ice nucleating proteins are classified according to biological origin, physical assembly of the cell membrane and specific genetic variants.
Source class classification identifies INPs found across organisms and how they utilize their protein for different ecological advantages. The bacterial class is the most studied and their main function is to facilitate precipitation or cause frost damage on plants causing various nutrients to release, the most common bacteria is the Pseudomonas syringae which is recognized as pathogenic to nature. The fungal class is a newly recognized class usually sharing bacterial ancestry but can have different structural arrangements. The plant source is used to facilitate water uptake or environmental interaction at sub zero temperatures. The insect class refers to certain freeze tolerant insects which naturally produce INPs in order to control where and when ice forms on in their bodies to prevent lethal intracellular freezing. INPs can also be classified according to activity type, or functional classes, categorized A, B and C according to the temperature at which ice nucleation is activated, this is directly tied to the size o the protein aggregate. Class A corresponds to the highly efficient INPs which nucleate ice between -2°C and -5°C, such as inaK, this type of nucleation requires very large and ordered protein aggregates and is usually connected to the presence of specific membrane lipids. Class B refers to moderately efficient INPs catalyzing ice between -7°C and -9°C, they appear as intermediate sized protein clusters. Class C identifies the less efficient INPs nucleating ice below -10°C, the activity is linked to smaller protein clusters or individual INP monomers. Finally, INPs can be classified according to the protein family and various species of bacteria contain different genetic orthologs of the ice nucleation genes. There is the Pseudomonas family, including inaZ, inaK, inaV, inaQ, they often share similar genetic structure such as N-terminal anchors, repetitive fragments and C-terminals. There is the Erwinia and Pantoea families originating from Gram-negative bacteria found in plants, including inaA, inaU and inaE. Additionally, as aforementioned there is the non-bacterial families such as the fungal or the insect families.
Choosing an INP for a project should depend on functional performance (its nucleation temperature capacity), the ease of expression and genetic stability, the membrane and lipid dependence, the stability of the INP and its environmental robustness, and, its safety and regulatory approval.
I am choosing to work with inaK as it offers more genetic stability, efficiency, reliability and it already has records of it being used in synthetic biology. Inak is frequently used as it has a superior compatibility with surface display, it is easier to anchor secondary protein to a cell surface using inaK over other INPs, the N-terminal of the inaK is highly optimized for integration into Gram-negative bacterial membranes and will be less likely to produce misfolds when anchoring. Additionally, inaK offers better fusion stability as it preserves its own folding and ice nucleating properties even if fused with larger fluorescent reporters such as sfGFP. Furthermore, inaK is a Class A INP with the ability to freeze at higher subzero temperatures it also offers better predictability and consistency as it has been more studied. According to Jung H.C. in his study ‘Expression of Candida antarctica lipase B on the surface of Escherichia coli using InaK anchoring motif’ , Enzyme and Microbial Technology, (1998), inaK’s N-terminal domain is a superior anchor for displaying functional enzymes on the surface of bacteria. Shi H. also demonstrates in his study ‘A novel surface display system using the ice nucleation protein InaK-N as an anchor for the directed evolution of a highly active organophosphorus hydrolase’ , Applied and Environmental Microbiology, (2015), that inaK’s anchor is very stable even in harsh environmental conditions, reinforcing the idea of using them for glacier blanket technologies. Moreover, Li Q. also highlights in his research ‘Surface display of Vitreoscilla hemoglobin on Escherichia coli using InaK-N and its effects on cell growth’, Letters in Applied Microbiology, (2009), that inaK can be combined with complex proteins while maintaining its ice nucleation functionality and the physiological health of the host cell. The codon optimized inaK sequence has already been well characterized in E. coli systems making it easy to use in synthetic biology. Furthermore, the study by Roeters in 2024 in The Journal of Physical Chemistry C shows inaK has a sensitivity to enhancers when maximum efficiency is needed, inaK can have the ability to undergo a 100-fold enhancement in the presence of compounds such as polyols as they can improve stability of the hydration order in the highly repetitive segments of inaK. Overall, inaK is the second INP to freeze at higher subzero temperatures after inaZ, both inaK and inaZ have been well characterized but inaK is more compatible with synthetic biology and it could easily be fused with an sfGFP, the inaK has extensively been studied. Inak is potentially the best choice amongst INPs as it responds very well to being modified and can be easily tailored according to what the end use is. In terms of working with safety regulations, working with Pseudomonas syringae would involve more difficulties as it is a classified pathogenic bacteria, however, Snowmax makes use of it in its inactive form which its safety has been EPA regulated. Additionally, the DNA sequence has already been recorded on UniProt meaning I can directly synthesize it using Benchling and Twist avoiding safety issues as the inaK protein is not pathogenic.
There are motifs necessary to INPs activity and function of ice nucleation
INPs share general features, here I will focus on the bacterial INPs. Bacterial INPs have a three domain structure and repetitive motifs. All bacterial INPs have a 𝞫-helix fold shaped by tandem repeats. INPs also all have exact spacing of Threonine and Serine residues matching ice lattice, this geometric spacing quality is a requirement for the protein to function as a template. The domain structure is always comprises a non repetitive N-terminal domain, (hydrophobic region which can anchor the protein to the outer membrane), a non repetitive C-terminal domain (hydrophilic tail assisting in protein stability and folding) and a central repetitive domain CRD ( the engine of the protein formed of tandem repeats which organize water molecules). The CRD is structured in hierarchy repeats composed of 16 amino acids which are then grouped in larger 48 residue periodicities. The entirety of the CRD is essential for ice nucleation at higher temperatures. Additionally, nucleation activity cannot occur without the 𝞫-helix fold meaning any mutations brought to the CRD has to be done in a way that it does not affect the 𝞫-helix fold outcome, this structurally essential. Moreover, the Threonine rich motif is essential to creating a water binding surface, if Threonine is replaced with non-polar residues then the protein might still fold correctly but risks losing nucleation capabilities, this is essential for functionality. Finally, the amount of repetition is essential as for instance a single 16 residue motif cannot nucleate ice, a certain mass of residue motif is required, explaining why inaK for instance has such a long repeat and demonstrates how it correlates to it being a very effective INP, this is essential for efficiency.
The effect of motif repetition should be considered and how repeat number affects nucleation activity.
Indeed, the idea that more repeats involve better or more nucleation is not so linear, the number of repeats defines which class the INP belongs to and how much nucleation it can produce. As aforementioned a 16 residue motif cannot produce any ice nucleation, there is a threshold of 15 to 20 repeats, which is the length of a typical CRD, to achieve the most basic level of ice nucleation at temperatures lower than -10°C. Below this number of repeats it is likely the protein could fold into a 𝞫-helix but it won’t catalyze ice as it will not have enough of a surface area to hold water molecules against temperature fluctuations. In order for the INP to function it needs an area large enough to stabilize ice catalyzation, this is referred to as the critical ice nucleus which is the smallest cluster of water molecules possible able to transform into a crystal rather than melt. The optimal number depends fully on the temperature the INP is supposed to be activated at or resist to. There is a correlation between the length of the sequence with which class the INP belongs to, the longer sequence belong to higher classes and vice versa. But, as much as longer repeats improve the capacity to nucleate at higher temperatures, too long of a sequence is likely to cause instability and become prone to genetic recombination and misfolding which can lead to failure to function. The activity does in fact reach a plateau as once the protein is long enough to stabilize a critical ice nucleus adding more repeats will not improve its function but rather might diminish its effectiveness, within each A B C class the INPs will plateau at their maximum capacity of repeats. Overcoming a plateau would depend more on aggregation capacity rather than length. The amount of repeats can impact aggregation, folding and membrane presentation. For class A the repeat number is essential to aggregation, longer repetitive sequences offer a better surface for proteins to stack on, without enough repeats proteins will not stack properly and won’t be very effective. The repeat number also impacts the folding process as the more the repeat number increases the larger the stress on the cell is for folding, the ribosome is challenged to reproduce with high fidelity a highly repetitive sequence. Finally, if the repetitive domain becomes too important it can become too heavy or too hydrophobic for the N-terminal anchor to be able to successfully connect the protein to the outer membrane, the protein might end up stuck in the cytoplasm where the INP function becomes useless.
INPs have specific mechanisms in order to organize water, template ice-like ordering and cluster ice nucleations at the membrane.
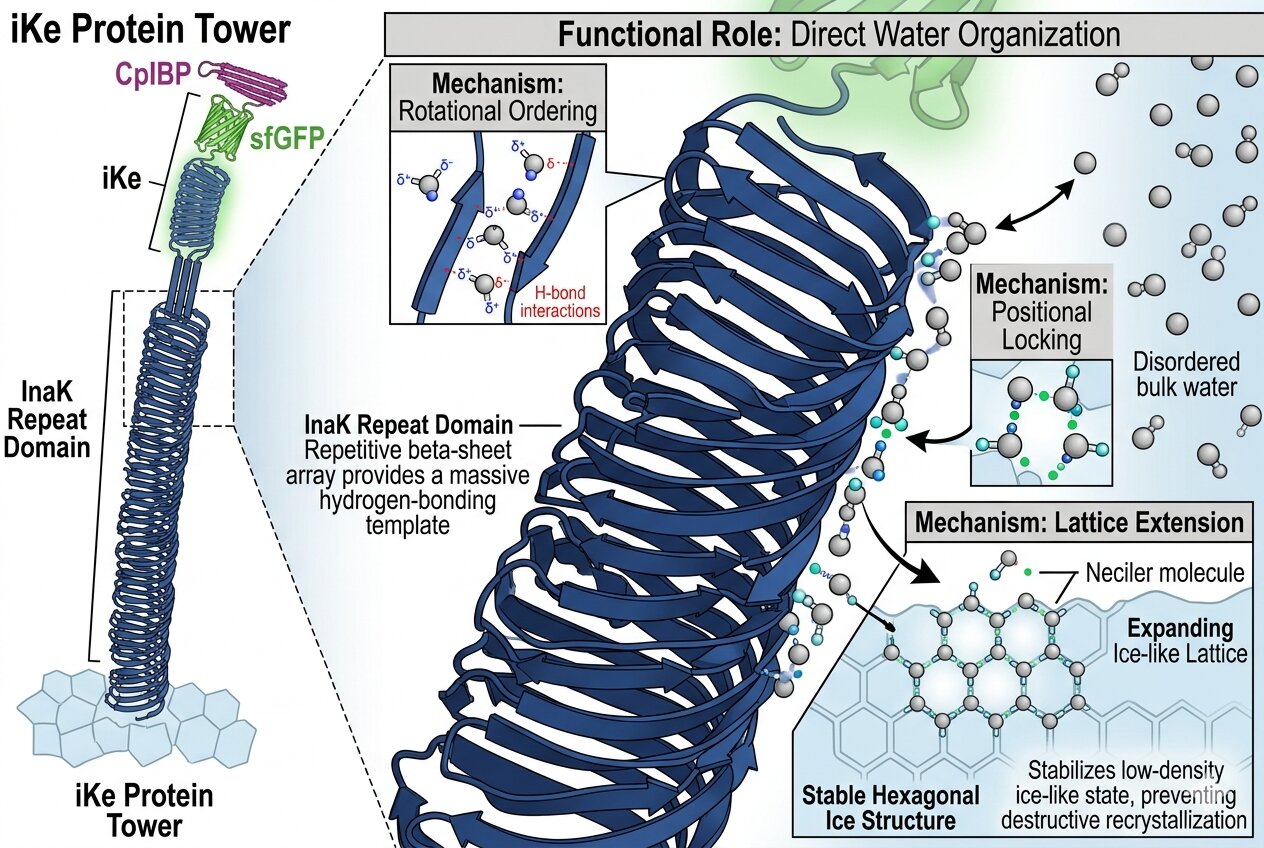
The molecular template and hierarchical assembly of INPs, especially inaK, able to organize water molecules into a structured solid crystal using only one protein is an incredible biological engineering mechanism of nature. An INP is able to actively organize water and direct its position through its hydrophobic and hydrophilic balanced motif. The 𝞫-helix structure provides a face with periodic motifs where the hydrophobic parts like Glycine prevent water molecules from attaching too strongly where in contrast the hydrophilic parts like Threonine groups form precise hydrogen bonds with the water molecules. This dual aspect of the surface offers a stable hydration layer as water molecules are linked to a 2D sheet imitating the surface of an ice crystal. The template ice-like ordering comes from the geometric matching of INPs’ structures with lattice matching, where spacing and distance of residues is very precisely organized in the repeat sections. Having an extremely organized and repetitive template allows the protein to use less energy and increase its nucleating ability as it facilitates the organization of the water molecules. Oligomerization or clustering affects activity by making it stronger or weaker. An individual INP is too weak to actively form ice whereas the class A INPs which are constituted of clusters have demonstrated to be more powerful, there is a correlation between the size of the repetitive section and the amount of ice the protein is able to produce. Additionally, a wider surface area can better stabilize a larger critical ice nucleus, which itself will be more resistant to higher temperatures making the INP more effective and stronger, this hierarchy is shown by Hudait in ‘Hierarchical assembly and environmental enhancement of bacterial ice nucleators’, Proceedings of the National Academy of Sciences, 2024. Membrane localization matters because for INPs to function correctly and effectively they must be situated in the cell membrane, therefore, even through a cell free design one would still have to synthesize a cell membrane. If the INP doesn’t reach the cell membrane (thanks to the N-terminal) then it is rendered useless, for ice catalyzation to be used it needs to occur on the outside of the cell.
INP production has progressed from harvesting wild type bacteria to synthetically engineered ones.
The native microbial production involved cultivating naturally ice nucleating bacterias such as the Pseudomonas syringae. The bacterias would be cultivated in large scale fermenters, once the desired density was reached the cell often deactivated through UV or chemical treatment to prevent environment damage from their pathogenic nature. Companies like Snomax use pelletized inactive Pseudomonas syringae. Recombinant in vivo bacterial expression is safer, non pathogenic, process of inserting the INP gene into a lab strain such as E.coli or B.subtilis, the protein will be expressed in the cytoplasm or led to the outer membrane. Cell free protein synthesis is used to produce INPs from DNA templates and added to a cell free solution, this allows to bypass the need for living cells and only requires the mechanical components such as the ribosomes, enzymes and amino acids extracted from a cell. This method is faster and offers better control for longer, repetitive proteins. Lastly, membrane based reconstitution or display systems are methods which combine recombinant expression with artificial membranes. For example, proteoliposomes is when INPs are retracted and reconstituted in a synthetic lipid vesicle, a liposome. This is the current method used to synthetically produce snow and allows researchers such as Hudait to study the different lipid types and understand the clustering required for high temperature activity.
INPs are now used in a wide range of applications from environmental and ecological focused uses and research to commercial sectors. These proteins are utilized in many industries as they can accurately control the ice nucleation phase.
As aforementioned, they are often used in artificial snow production, as demonstrated by Snomax International using native bacterial proteins for snow making at temperatures where traditional machines would not be effective. The use of INPs in snow production significantly reduces the energy and water cost for ski resorts and therefore reduces the commercial impact on the environment. This application is close to my area of research as it is used within a similar context of mountains and rebuilding and reinforcing skiing tracks against rising temperatures. However, the mindset and end use varies widely, the aim to protect and preserve glaciers does not have a commercial use as it focuses on an ecological solution to climate change rather than compensating for climate change consequences human activity does not want to face. Additionally, the technology would still vary as the aim for me is not to produce snow but ice which will have different temperature requirements and a different upkeep. My research of INPs would come closer to the geo-engineering experimental approach of considering the use of INPs for cloud seeding, where silver iodide is commonly used and can have some toxic secondary effects but INPs are biodegradable and highly efficient making them a possible sustainable alternative. These approaches are still conceptual and theoretical projects as seen in the following projects; for atmospherical cloud seeding in the Walser 2024 project ‘Fungal ice nucleation proteins open new pathways for weather modification and biopreservation’, Science Advances, or, in idea of developing glacier blankets in the case study of Biotreks 2021, ‘Ice nucleation proteins – a synthetic pathway to alleviate ice loss’ . INPs are also commonly used in the food industry as freezing structuring and preservation technologies allowing to control the size and distribution of ice crystals. Large ice crystals can damage the texture of frozen food whereas precise ice nucleation can preserve it, the INPs are used as freeze structuring agents to form many small ice crystals simultaneously. Similarly to the artificial snow making technologies using INPs to freeze food also allows to reduce energy waste as the process is controlled and optimized. Furthermore, INPs are used in the biotechnology or medical field for biopreservation. INPs have the ability to prevent supercooling and are used to better preserve sensitive biological samples. For example it can be used for cryopreservation where samples are frozen in a homogeneous and controlled way preventing cells from being damaged or dying. There are case studies where this technology is being experimented with in organ preservation, with controlled nucleation in liver preservation as discussed in ‘Controlled ice nucleation by ice-nucleating proteins for the cryopreservation of complex biological systems’, Biomaterials (38, pp. 11–21. doi: 10.1016/j. ), 2015, by Lee, C.Y., et al..
For this section I made use of Gemini assistant by inputting my information collected with the comparative elements needed so I could create a clear comparative table.
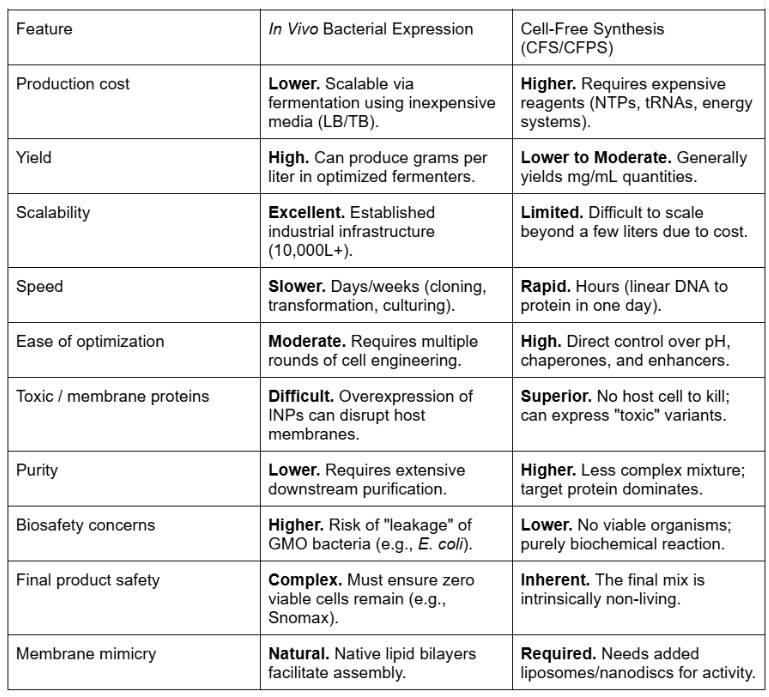
There are clearly very strong advantages to using CFPS compared to an in vivo method. My main challenge when working with inaK, a class A INP, will be the membrane dependence issue, as class A INPs require a membrane in order to form larger ice clusters using the lipid bilayer. Class C INPs would be able to function without a membrane but would be significantly less useful for the needed output working on glacier preservation faced with the rise of temperatures as they will produce much less ice and much lower sub zero temperature. An in vivo method using bacteria naturally provides a base for the protein to attach to, however, large amounts of inaK can become toxic to the host and might just become ineffective. Moreover, CFPS offers better optimization and enhancement enabling a superior platform of the 100 fold provided by polyols. In a cell free design the polyol concentration can be precisely adjusted to avoid killing the host which can enable better maximization to template inaK. The other main issue I would be faced with using CFPS is the limited scalability as it can be more costly, using an in vivo method the INP like inaK could be inoculated into a glacier in a new host and naturally form horizontal gene transfer, which would not be possible with CFPS. Finally, CFPS has much less regulatory and biosafety issues which will be extremely relevant for my project as my technology would be directly inoculated into nature and the natural consequence this could have must be very carefully considered.
Understanding inaK and INPs involves understanding whether using the whole protein or a fragment is the most effective. Interfering with the natural structure of an INP, here inaK, can completely shift its ability to correctly nucleate ice.
Expressing the full protein allows for the hierarchical architecture which focuses on geometry. The N-terminal and C-terminal cannot be taken out of the structure and the first plays the essential role of an anchor and the second is essential in the folding process, with these the central repetitive domain would likely fail to nucleate ice, its primary function. Additionally, the large size of the inaK gives it stability and ensures the protein can withstand the mechanical stress it undergoes in a synthetic cell surface or within a Glacier Blanket concept for instance, it is “important to strike a balance between adequately mutating conserved residues and avoiding large-scale disruptions to the overall fold of the repetitive region”, (Forbes, 2022). As aforementioned a single nucleation motif cannot function alone, a single motif would be too small to overcome the thermal energy of liquid water, the long continuous and repetitive surface of the protein is essential to stabilize the critical ice nucleus. Additionally the 𝜷-helix is a critical part of the structure as this is what fold the individual strings of amino acid into the 𝜷-helix shape again essential to stability and the nucleation of ice. A truncate construct would on one hand reduce the genetic burden during PRC or translation as the sequence would be shorter and could offer higher reliability in data recovery. However, on the other hand, as aforementioned this method would simply reduce the ice nucleation capability and efficiency and essentially would downgrade the protein, likely reducing an inaK protein from a class A to a class C. Again, removing the scaffold would render the INP useless as without the N-terminal anchor it will not be able to orient itself to the membrane and might be able to template a few water molecules but won’t be able to form crystal lattice as it will not have a flat 2D surface and the motif will randomly float within the cell. Additionally, the N-terminal anchor is also what enables clustering, without it the protein will not efficiently nucleate ice. Thus, the motifs provide the chemical code of the hydrogen bonding patterns, the spacing and repetition provides the physical geometrical template allowing the ice to form, and, the supramolecular assembly provides the scale neede for a class A INP like inaK to nucleate ice at a higher temperature.
The membrane is an essential part of an INPs ability to express ice nucleation.
The membrane is not simply a container for the protein, in this case the membrane becomes a functional factor or tool. Ice nucleation produced by an INP like inaK forms on the surface of the cell at the membrane for better more effective ice catalyzation. Without a membrane imitating scaffold like a liposome or nanodisc the INP, inaK, would fail to reach its class A efficiency. The membrane is essential for clustering, a lipid bilayer would act as a fluid 2D scaffold and the inaK would be restricted to a 2D plane which increases concentration. Orientation is not possible without a membrane, the protein would simply tumble in the space and not align to each other preventing ice lattice, the N-terminal ensures proper orientation with the membrane. On a mechanical aspect the membrane offers a resilient yet flexible scaffold preventing the template from collapsing. A membrane facilitates multivalent display as it becomes a hub for ice binding sites to form simultaneously. The membrane also plays a critical role in the protein folding as the lipid tales help the N-terminal domain to work properly in a hydrophobic environment. In CFPS system the lipid membrane is vital to avoid dead aggregates, inclusion bodies, and give the protein a direction upon translation. Overall, a membrane of membrane like material is essential for ice nucleation and makes it more stable and efficient. For my project I will need to create a synthetic cell membrane.
Understanding the literature gaps allows to better see the potential and limitations within research and development .
While we know how to identify INPs and how their general mechanism works there are still many existing gaps within literature. For instance, while there is an understanding of the different classes and that 8.33 MDa didecamers correlates with high temperature nucleation there is still uncertainty around the tipping point of minimal clusters, of how many proteins are actually needed. Additionally, there is a lack of high resolution structural data mapping the transition from a single 𝞫-helix monomer to a supramolecular assembly and without understanding what is the minimal viable cluster it makes it difficult to optimize synthetic cells the best efficiency with the least protein expression. On top of it, the relationship between repetitive sequence length and the freezing temperature is still to some extent speculative as INPs are usually tested on within similar context using the same amount of repeats and working within a membrane like environment, as explorations have been limited it is likely a lot of potential has not yet been explored. Moreover, the main evident gap in literature is the lack of knowledge on how to achieve class A function without relying on a membrane or membrane like environment. There is also limited information on the comparative data between in vivo and CFPS systems, currently it appears to be more of an overall idea. Finally, the potential of INPs has not been explored much further from what we know already, the idea of tailored design through the exploration of specific mutations has not been explored. In conclusion, in the context of my glacier preservation project using inaK I am faced with a few literature gaps regarding clustering requirements, precise manufacturing comparison and varied environmental application.
I believe from this research that the best current option to explore relies around protein engineering and chimeric design. Structural modifications appear less likely to be successful from research.
My personal interrogation to redefine and push my aims:
Can I increase the efficiency of inaK by combining it with another compound?
sfGFP tag : increased stability and UV protection with both aspects increase ability to produce ice nucleation and be more resilient to higher temperatures (whether increasing the nucleation point or afterwards resisting warmer external temperatures)
Polyol : (Sorbitol, Glycerol, Xylitol) a hydroxyl rich molecule which can behave as a partner template, it cannot nucleate ice itself but can help organize the water molecules around the inaK repeats reducing potential entropy
Ions : adding specific ions to a CFPS design can stabilize the 𝞫-helix and inaK repeats improving nucleation
Ice Binding Domains (IBDs): added to the C-terminal help the INP adhere to the surface of another layer of ice (such as the glacier) preventing INPs to be naturally washed away by meltwater
Hydrophins : small surface active protein enable INP to spread evenly across a surface ( such as the ice layer of a glacier)
Melanin / Scytonemin : pigments which could serve as a sunscreen to the INP, protecting it from UVs would make it more resistant to melting
Lipid nanodiscs/liposomes: in CFPS combining inaK with a synthetic scaffold will increase the productivity of the N-terminal anchoring the INP with a stronger bond to the membrane
Protein cage: is a method where the INP is fused to self assembling protein cage (such as Ferritin or Encapsulin) combining inaK proteins into a larger single molecule creating a super-cluster by design rather than relying on the membrane activity
Reference list
Arnold, D.L. and Preston, G.M. (2019). Pseudomonas syringae: enterprising epiphyte and stealthy parasite. Microbiology, 165(3), pp.251–253. doi:https://doi.org/10.1099/mic.0.000715. Boztas, S. (2024). Pumped up: will a Dutch startup’s plan to restore Arctic sea-ice work? The Guardian. [online] 27 Feb. Available at: https://www.theguardian.com/environment/2024/feb/27/climate-crisis-arctic-ecosystems-environment-startup-plan-pump-restore-melting-sea-ice-caps. Biotreks (2021) ‘Ice nucleation proteins – a synthetic pathway to alleviate ice loss’, Biotreks, (e202111).
Christner, B.C., Morris, C.E., Foreman, C.M., Cai, R. and Sands, D.C. (2008). Ubiquity of Biological Ice Nucleators in Snowfall. Science, 319(5867), pp.1214–1214. doi:https://doi.org/10.1126/science.1149757. Davies, P.L. (2014) ‘Ice-binding proteins: a remarkable capacity to adapt for life at cold temperatures’, Biochemical Journal, 458(1), pp. 9–20. doi: 10.1042/BJ20131291.
Dr. Tobias Weidner, Dr. Janine Fröhlich-Nowoisky (2016). The effect of bacterial ice nuclei. [online] Www.mpg.de. Available at: https://www.mpg.de/10470442/ice-formation-bacteria-syringae. Experimental Data Figure (2026) Intact Mass Spectra: Native vs. Denatured sfGFP Analysis.
Experimental Data Figure (2026) Sequence Coverage Report: 88% Confirmation of sfGFP.
Experimental Data Figure (2026) CDMS Spectrum of KLH Oligomeric States.
Forbes, J., Bissoyi, A., Eickhoff, L., et al. (2022) ‘Water-organizing motif continuity is critical for potent ice nucleation protein activity’, Nature Communications
Garnham, C.P. et al. (2011) ‘A conserved water-organizing motif in ice-nucleating proteins’, Molecular Microbiology, 79(6), pp. 1419–1427. doi: 10.1111/j.1365-2958.2011.07546.x.
GoldBio (n.d.) The Extraordinary Bacterial Proteins That Make Snow. Available at: https://www.goldbio.com/blogs/articles/the-extraordinary-bacterial-proteins-that-make-snow (Accessed: 25 April 2026).
Govindarajan, A.G. and Lindow, S.E. (1988) ‘Size of bacterial ice-nucleation sites measured in situ by radiation inactivation’, Proceedings of the National Academy of Sciences (PNAS), 85(5), pp. 1334–1338.
Hudait, A. et al. (2024) ‘Hierarchical assembly and environmental enhancement of bacterial ice nucleators’, Proceedings of the National Academy of Sciences (PNAS), 121(18), p. E2409283121.
James Dalton, Global Head, Water and Wetlands Team, IUCN (2025). Protecting glaciers – our most effective natural water manager. [online] IUCN. Available at: https://iucn.org/blog/202503/protecting-glaciers-our-most-effective-natural-water-manager. Jung, H.C. et al. (1998) ‘Expression of Candida antarctica lipase B on the surface of Escherichia coli using InaK anchoring motif’, Enzyme and Microbial Technology, 22(5), pp. 348–354.
Lee, C.Y., et al. (2015) ‘Controlled ice nucleation by ice-nucleating proteins for the cryopreservation of complex biological systems’, Biomaterials, 38, pp. 11–21. doi: 10.1016/j.biomaterials.2014.10.050.
Li, Q. et al. (2009) ‘Surface display of Vitreoscilla hemoglobin on Escherichia coli using InaK-N and its effects on cell growth’, Letters in Applied Microbiology, 49(1), pp. 71–76.
Ling, M. L. et al. (2018) ‘The constructive role of protein repeats in ice nucleation’, Nature Communications, 9, p. 3314.
Lukas, M. et al. (2025) ‘A New Class of Fungal Ice-Nucleating Proteins with Bacterial Ancestry’, ChemRxiv. doi: 10.26434/chemrxiv-2025-73058.
Lindow, S.E. et al. (1989) ‘Relationship between Ice Nucleation Frequency and inaZ Protein Content in Escherichia coli’, Molecular Plant-Microbe Interactions, 2(5), pp. 262–272.
McDonough, F. (n.d.). What is Cloud Seeding? [online] Desert Research Institute. Available at: https://www.dri.edu/cloud-seeding-program/what-is-cloud-seeding/.
O’Sullivan, D. et al. (2016) ‘The influence of pH, ionic strength and soluble organics on the ice nucleating ability of Pseudomonas syringae’, Atmospheric Chemistry and Physics, 16(11), pp. 7443–7454
Pandey, R. et al. (2016) ‘Ice-nucleating bacteria control the order and dynamics of interfacial water’, Science Advances, 2(4), p. e1501630. doi: 10.1126/sciadv.1501630.
Ponte di Legno Tonale. (n.d.). The protection of Presena Glacier. [online] Available at: https://www.pontedilegnotonale.com/en/pontedilegno-tonale-what-to-see/the-protection-of-presena-glacier/. Roeters, S. J. et al. (2024) ‘Polyol-Induced 100-Fold Enhancement of Bacterial Ice Nucleation Efficiency’, The Journal of Physical Chemistry C, 128(15).
Roeters, S.J., Golbek, T.W., Bregnhøj, M., Drace, T., Alamdari, S., Roseboom, W., Kramer, G., Šantl-Temkiv, T., Finster, K., Pfaendtner, J., Woutersen, S., Boesen, T. and Weidner, T. (2021). Ice-nucleating proteins are activated by low temperatures to control the structure of interfacial water. Nature Communications, [online] 12(1), p.1183. doi:https://doi.org/10.1038/s41467-021-21349-3.
Schmid, D. (2026) Glacier Blankets Could Help Prevent Melting. [Online Video]. 25 April. Available at: https://www.youtube.com/watch?v=hKT_SGK2qtY (Accessed: 27 April 2026).
Schoborg, J.A. et al. (2014) ‘Aqueous two-phase system (ATPS) for direct fractionation of proteins from cell-free protein synthesis’, Biotechnology and Bioengineering, 111(12), pp. 2405–2415.
Silverman, A.D. et al. (2020) ‘Cell-free gene expression: an expanded repertoire of applications’, Nature Reviews Genetics, 21(3), pp. 151–170. doi: 10.1038/s41576-019-0186-3.
Shi, H. et al. (2015) ‘A novel surface display system using the ice nucleation protein InaK-N as an anchor for the directed evolution of a highly active organophosphorus hydrolase’, Applied and Environmental Microbiology, 81(15), pp. 5128–5135.
Snomax.com. (2015). FAQ - Snomax. [online] Available at: https://www.snomax.com/faq.html. Snomax International (2026) The Science of Snomax: Maximizing Snow Production Efficiency. [Online Technical Bulletin].
Steroplast Healthcare (2022). How Do Instant Ice Packs Work? [online] www.steroplast.co.uk. Available at: https://www.steroplast.co.uk/knowledge-base/how-do-ice-packs-work.html. Un-glaciers.org. (2026). Glacier Preservation is the Key to Ensuring the Security of Water, Energy, and Environmental Resources. [online] Available at: https://www.un-glaciers.org/en/articles/glacier-preservation-key-ensuring-security-water-energy-and-environmental-resources. USNSJ (n.d.) ‘Wonders of the Invisible World: Pseudomonas syringae, the Ice Maker’, University of Southern North Science Journal, 2(2).
Wadokar, T. (2025). Snow Seeds - Tanay Wadodkar - UAL Showcase. [online] Arts.ac.uk. Available at: https://ualshowcase.arts.ac.uk/project/635735/cover [Accessed 9 Feb. 2026].
Walser, A. et al. (2024) ‘Fungal ice nucleation proteins open new pathways for weather modification and biopreservation’, Science Advances, 10(12), p. eadl1234. doi: 10.1126/sciadv.adl1234.
Wellpott, V. and Wellpott, V. (2025). Glacier protection with geotextiles – A sustainable solution for the future. [online] Naue - Geosynthetics | Digtal Engineering Software | Installation services. Available at: https://www.naue.com/glacier-protection-with-geotextiles-a-sustainable-solution-for-the-future/.
Why does this project matter?
Addressing the issue of glaciers melting means directly confronting climate change. It might seem like it but glaciers protect the Earth and its ecosystems which we are an active part of by helping to regulate the Earth’s temperature, with the deregulation of temperature comes a rise of temperatures and a disruption of the natural cycle of melting and reforming of glaciers. With increasing temperatures glaciers melt more intensely in spring and summer, less snow is produced during winter and winters are not cold enough to correctly and efficiently rebuild enough layers of ice. The weaker glaciers grow as years pass the less resistant they become to warmer temperatures and as their surface diminishes the less they are able to protect ecosystems. While some attempts are done to protect glaciers I argue that not nearly enough is being done most likely because protecting glaciers does not directly create a financial profit, less investments are being made in research projects aiming to protect nature (which directly benefits the health of human and non human species) compared to human centric research. This project demonstrates how we could make use of existing research and projects studying and using INPs and optimize and push further this knowledge to shape an innovative solution to glaciers melting. The aim of this project and the DNA construct outcome is to experiment with how to increase the activity and function of natural mechanisms such as ice nucleation without distorting completely how nature already performs it. This design offers a supportive system respectful of what nature has already created rather than a core modification into something unnatural. This project can highlight an issue too often overlooked as well as providing more data about glaciers, as they would have to be thoroughly studied in order to work with them, and the inaK INP which my project focuses on, an INP studied only to an extent. This research would add a large amount of qualitative and quantitative data, observations and explore in depth the potential of a class A INP which might open up new exploratory routes and inspire more bioremediation projects.
Ethics Context
Working and researching on a delicate, threatened ecosystem is no mundane task, it requires care and vigilance. No synthetic biology project which may involve (genetic) modifications of an environment should be done lightly, synthetic biology is a powerful tool and has the capacity to be very beneficial or detrimental to a system. Ethical considerations of this project are vital to the well being of natural ecosystems.
Geopolitics play a large role in how glaciers are treated. As the Geneva Environment Network states we “ require targeted policy and a geopolitics of ice” and “this environmental issue is directly linked to human rights” (“Unprecedented Rates of Mountain Glacier Melting | Glaciers and the Role of Geneva”). Laws and objectives have been progressively put in place with a goal of glacier protection and tracking. Firstly, the UN has consistently raised awareness for the need of national initiative to protect glaciers, such as during the coming seventh United Nations Environmental Assembly in December of 2026. The UNEP is in agreement with the UN’s Decade on Ecosystem Restoration calling to prevent and reverse any harm brought to mountain ecosystems. This was taken further under the Adaptation at Altitude programme where UNEP is a partner to the Swiss Agency for Development and Cooperation where the goal was to find innovative mountain solutions around the world in order to regroup policy representatives from a variety of countries in order to share plans surrounding mountain and glacier preservation solutions. Moreover, the High Mountain Summit of October 2019 held in Geneva called for actions with the goal “to support more sustainable development, disaster risk reduction and climate change adaptation” (“Unprecedented Rates of Mountain Glacier Melting | Glaciers and the Role of Geneva”). Additionally, global data collection about glacier evolution started in August of 1894 with the foundation of the International Glacier Commission during the sixth International Geological Congress in Zurich. On an Italian scale, for the purpose of this case study, there is a lack of a singular law protecting glaciers, an aspect within this project which could aim to change through petitioning and lobbying for policy change, synthetic biology needs to extend further than the research and development. However, Italy has made mountain glaciers public property preventing any private appropriation of natural resources such as water, plus regional committees in conjunction with the Italian Alpine Club and other specialized committees oversee high altitude areas in order to prevent unsustainable development. Overall, there are wider legal and policy considerations to understand and respect in order to research and work in a protected natural environment.
Permits, working and researching on glaciers requires strict permits which I would obtain in order to further this project. Permits (here in the case of my local inspiration of the Italian Alps) include authorizations from National Protected Parks, the Stelvio National Park. As well as regional authorisations to access and research local glaciers, this includes permits for samples and equipment. Action would require a declaration to the Italian Glaciologue Committee (IGC). Ethical clearance would be needed by global institutions such as the Royal Geographical Society and local institutions like the Italian National Research Council ( Consiglio Nazionale delle Ricerche)
Furthermore, I would aim for transparency and clear communication, while specific technical knowledge might not be shared the overall impact of this projects, in how it is situated, its purpose, the way it is being done, the final output should always be shared globally and with the local communities as it directly affects nature and human health and lifestyle.
Team and expert knowledge, in the scope of this project and ensuring to the best of my abilities the ethical preservation of (mountain) glaciers I would work with a team of varied experts which I could rely on for knowledge and feedback. I am not a scientist, I am a designer and a local seeing first hand the disappearance of glaciers. Attempting to problem solve on my own would not be effective or successful. I would build a team of : glaciologists (as they have the knowledge of glaciers), synthetic biologists (as they have the required knowledge and skill to see this project to term), conservation biologist ( as they have the aim and knowledge to protect biodiversities, they need to be familiar with the local biodiversity), a botanical biologist (as they would have a wider perspective of the impact of this work on the surrounding flora), a geologist (familiar with the local environment, again to better understand the area and the effects of this project), a textile engineer (with the practical design knowledge for aim 3), a biodesigner (myself with a systems thinking and innovative design approach) and last but not least, the park rangers (locals with direct knowledge of the environments, usually protected, with a social cultural historical attachment to the land). Moreover, I would aim to collaborate with local university departments and public institutions such as the Consiglio Nazionale delle Ricerche also specialized in local biodiversity or glaciology in order to expand further input knowledge and resources and reinforce the success of outcome. The aim of building a team with varied profiles is to anticipate any harmful consequences this project could have on the environment. Any small change or addition to a functioning ecosystem can have drastic consequences even if a project, technology or species wouldn’t appear harmful at first. This project should not be put to use on glaciers until it has been verified thoroughly that it could only benefit glaciers and the surrounding ecosystems.
Reference List
Anacona, P.I., Kinney, J., Schaefer, M., Harrison, S., Wilson, R., Segovia, A., Mazzorana, B., Guerra, F., Farías, D., Reynolds, J.M. and Glasser, N.F. (2018). Glacier protection laws: Potential conflicts in managing glacial hazards and adapting to climate change. Ambio, 47(8), pp.835–845. doi:https://doi.org/10.1007/s13280-018-1043-x.
Genevaenvironmentnetwork.org. (2024). Unprecedented Rates of Mountain Glacier Melting | Glaciers and the Role of Geneva. [online] Available at: https://www.genevaenvironmentnetwork.org/fr/ressources/nouvelles/unprecedented-rates-of-mountain-glacier-melting/.
ISPI. (2024). Mr. Matterhorn and Its Glaciers – A Critical Exploration of the Protection of Glaciers through Rights of Nature | ISPI. [online] Available at: https://www.ispionline.it/en/publication/mr-matterhorn-and-its-glaciers-a-critical-exploration-of-the-protection-of-glaciers-through-rights-of-nature-187115 [Accessed 13 May 2026].
NANGERONI, G. and VANNI, M. (1963). THE ACTIVITIES OF THE ITALIAN GLACIOLOGICAL COMMITTEE ON THE OCCASION OF THE INTERNATIONAL GEOPHYSICAL YEAR (IGY) / L’activité du Comité Glaciologique Italien à l’occasion de l’Année Géophysique Internationale (A. G. I.). International Association of Scientific Hydrology. Bulletin, 8(3), pp.97–101. doi:https://doi.org/10.1080/02626666309493342.
Rgs.org. (2026). RGS Explore Grants. [online] Available at: https://www.rgs.org/exploration/grants/expedition-grants/rgs-explore-grants [Accessed 12 May 2026].
UN (2021). UN Decade on Restoration. [online] UN Decade on Restoration. Available at: https://www.decadeonrestoration.org/. www.cnr.it. (n.d.). Home | Consiglio Nazionale delle Ricerche. [online] Available at: https://www.cnr.it/en.
Design Development
Final Design
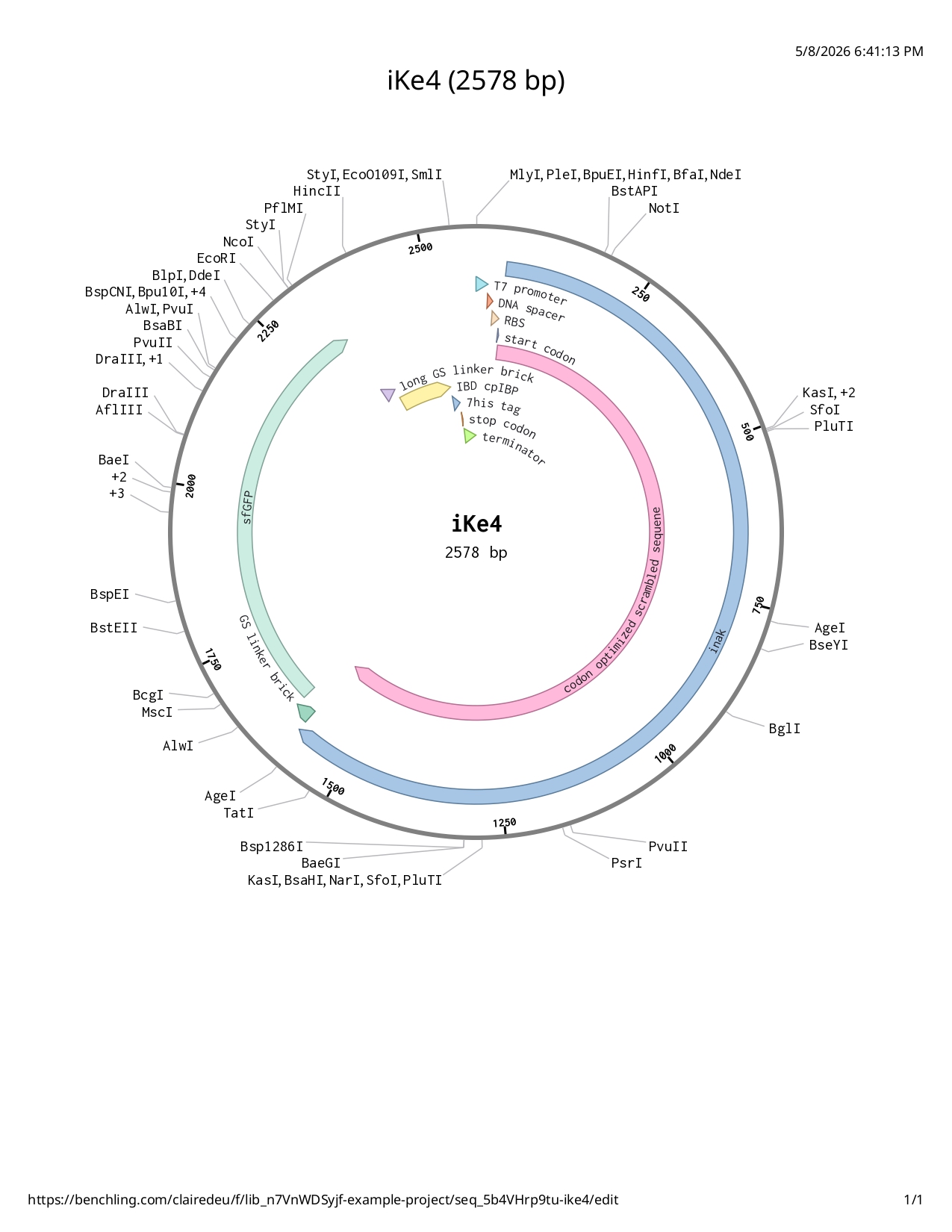
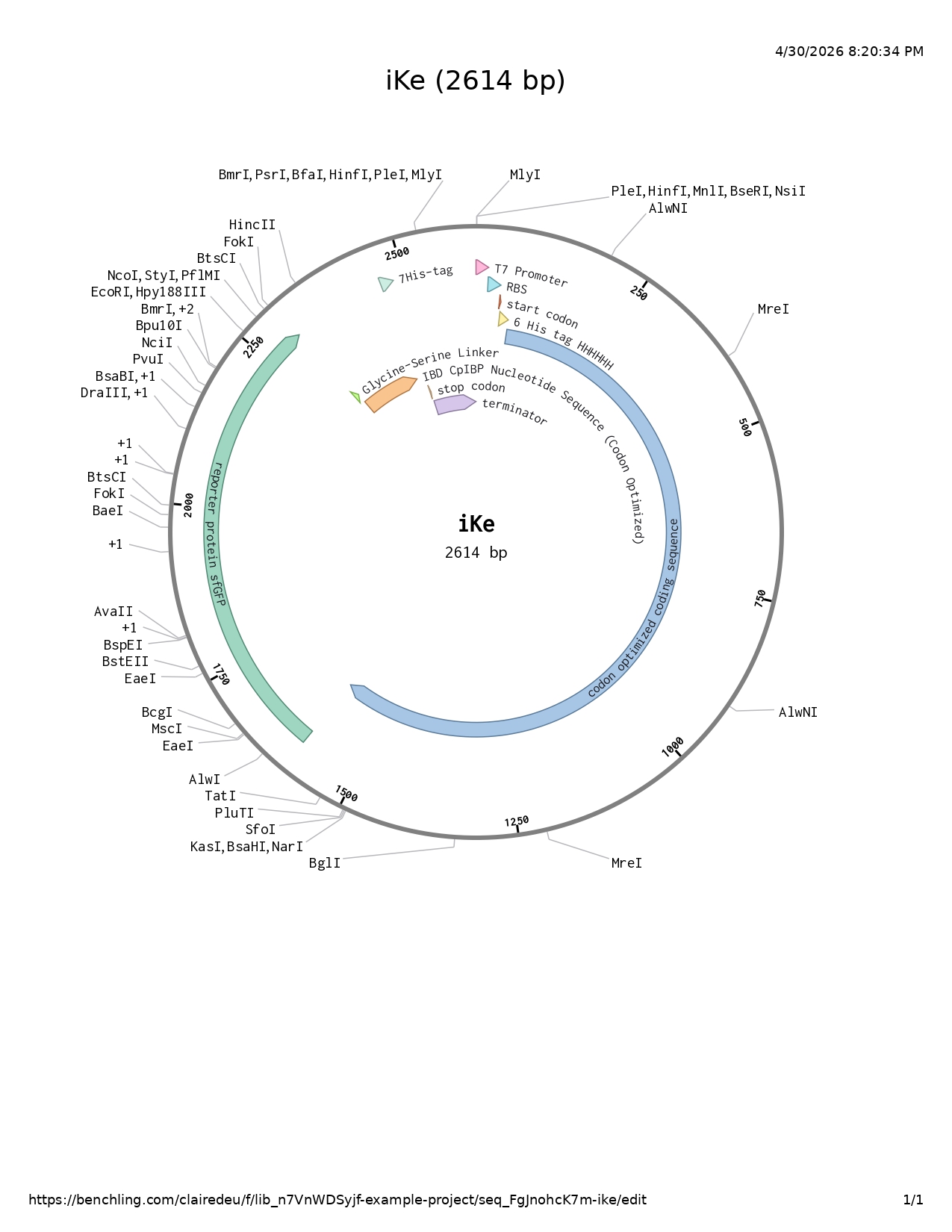
- T7 promoter
- RBS
- Start codon
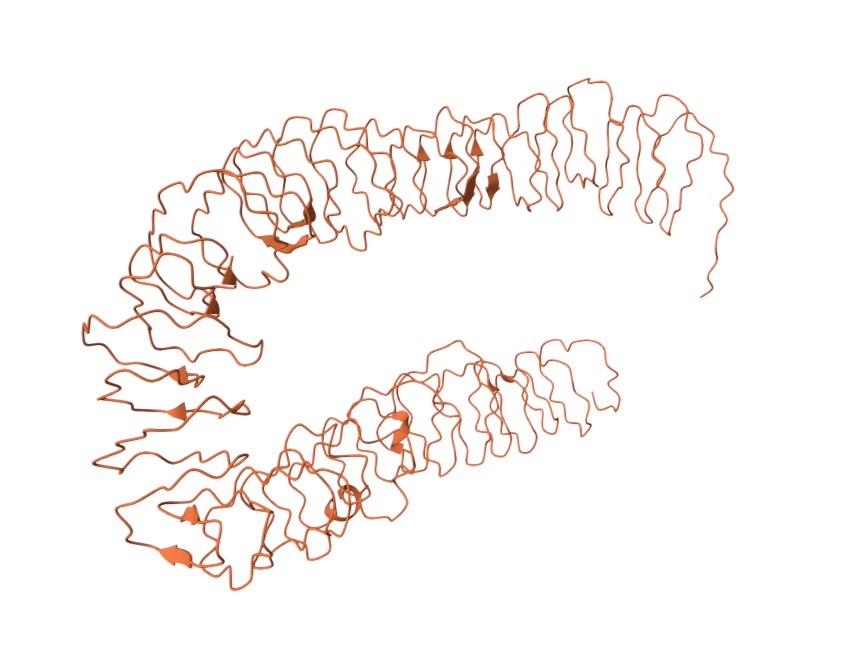
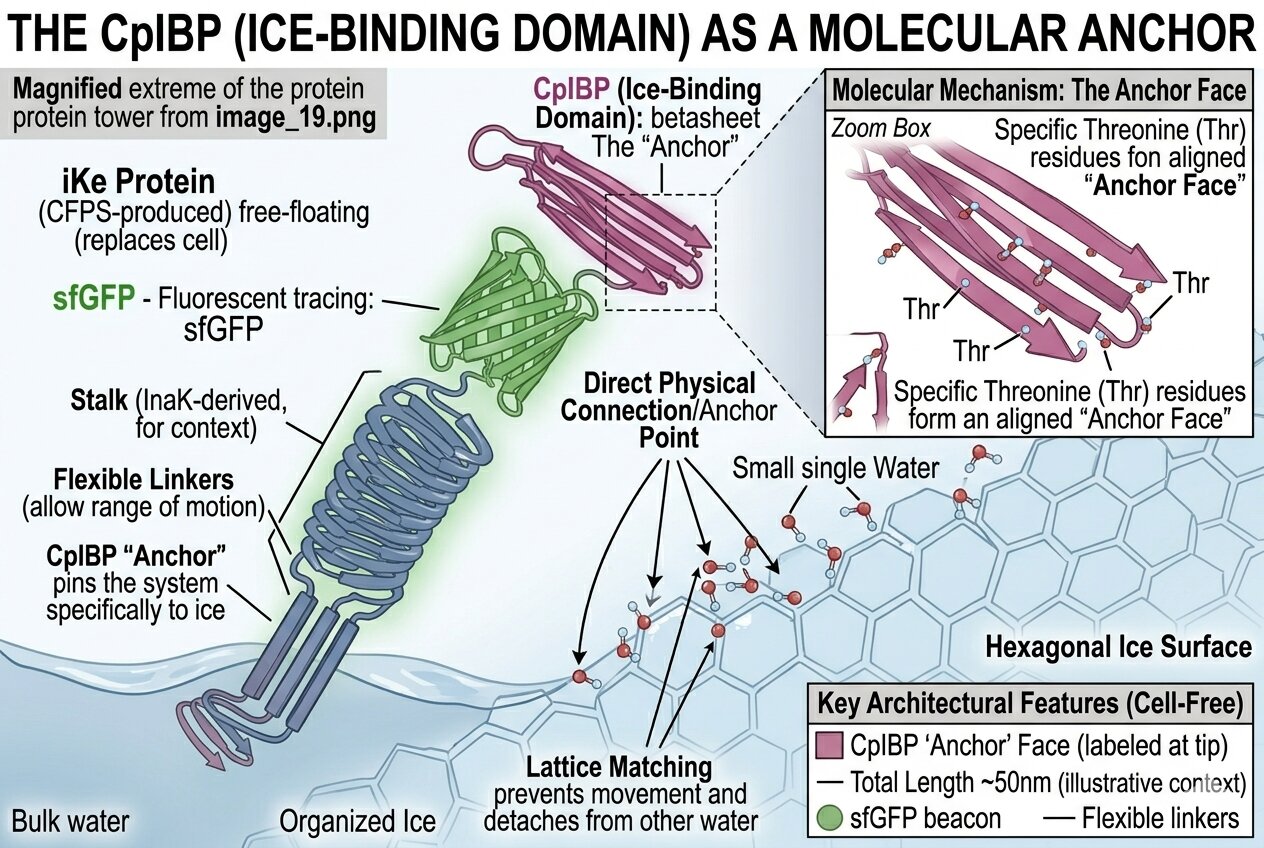
- Codon optimized and scrambled coding sequence of inaK: offers class A nucleation (scrambling the sequene allowed me to limit repetitive segments in the sequence)
- Reporter protein sfGFP: increased stability and UV protection with both aspects increase ability to produce ice nucleation and be more resilient to higher temperatures (whether increasing the nucleation point or afterwards resisting warmer external temperatures)
- Glycine serine linker (GGGGSGGGGS): A flexible tether that allows the IBD to move freely and “find” the ice surface.
- IBD codon optimized: added to the C-terminal help the INP adhere to the surface of another layer of ice (such as the glacier) preventing INPs to be naturally washed away by meltwater with CpIBP: The anchor that locks the entire complex to the glacier.
- 7His-tag : protein purification very suitable with the sfGFP
- Stop codon
- Terminator
I used Pymol to visualize the inaK protein (unmodified strain) I worked on

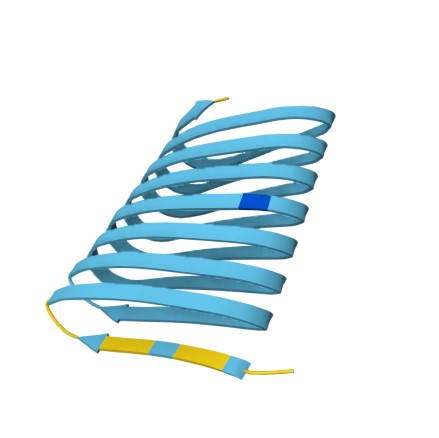
I used Alpha Fold 3 to visualize the sfGFP and IBD I chose to add to my protein sequence
sfGFP protein reporter
Ice binding domain
How my iKe construct works:
In order to visualize what was happening on a molecular level I generated the following illustration using Gemini
How is the construct interacting with its environment?
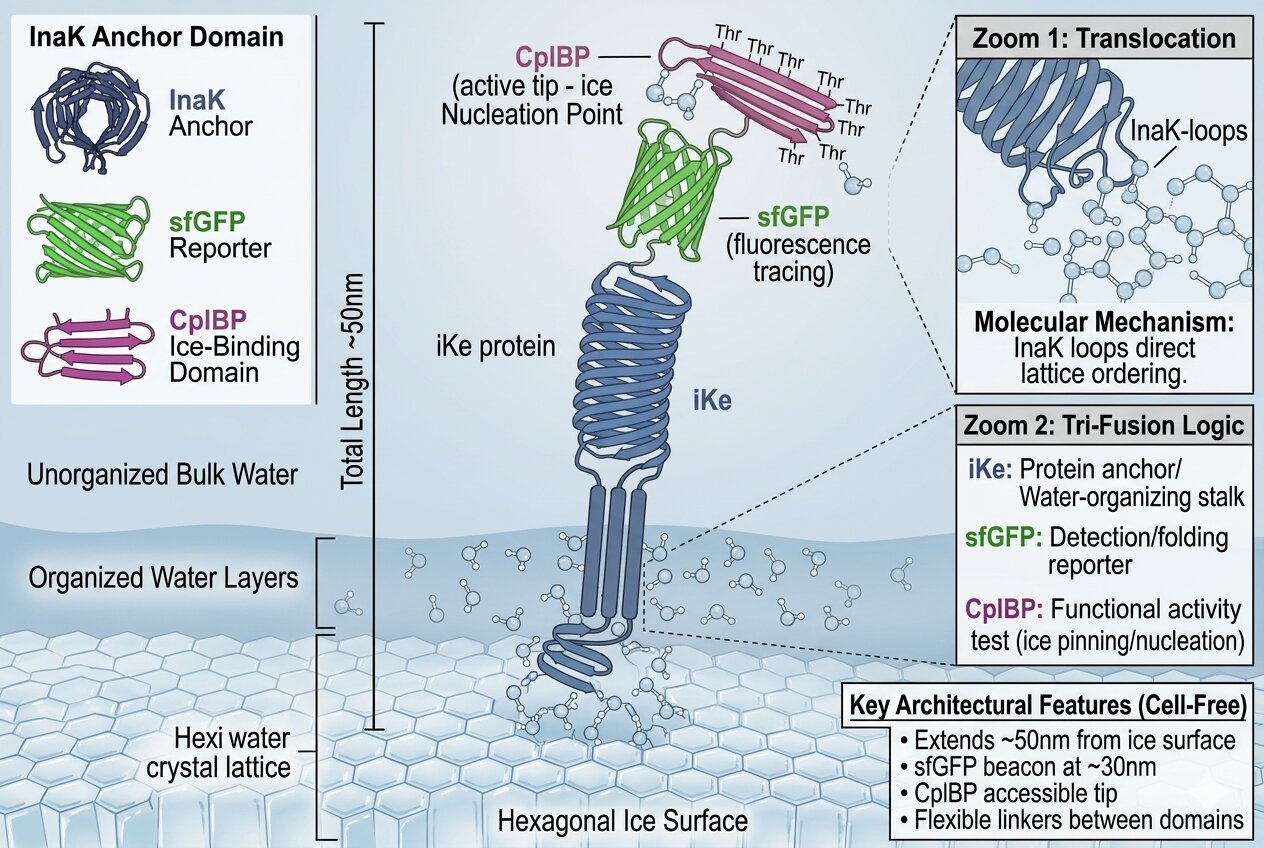
How is the protein organizing water molecules?
How is the ice binding domain IBD connecting to the ice surface of the glacier?
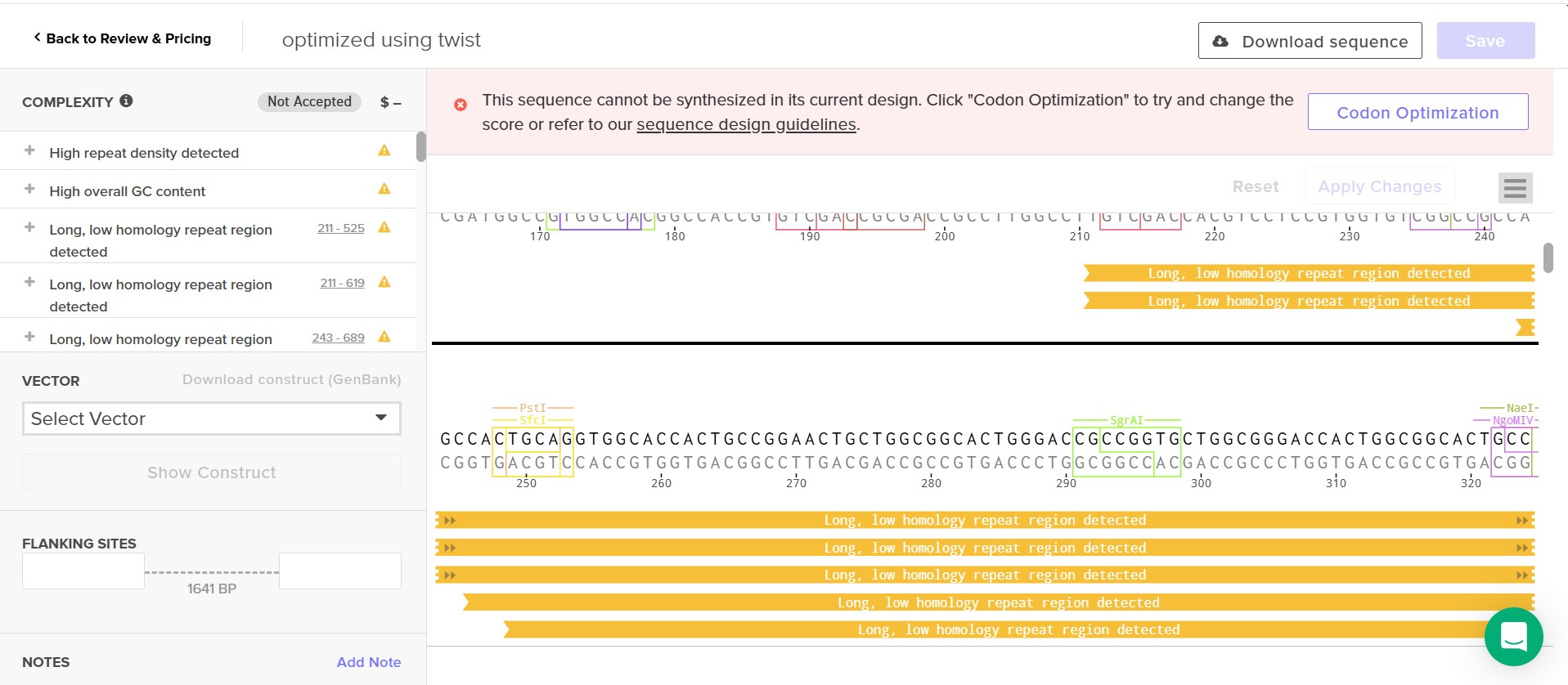
As you will see from the translation of my sequence and the explanation in the design development process using Twist to order my sequence appears challenging, my construct is and needs to be long and repetitive which presents too much complexity for Twist even after scrambling my sequence.
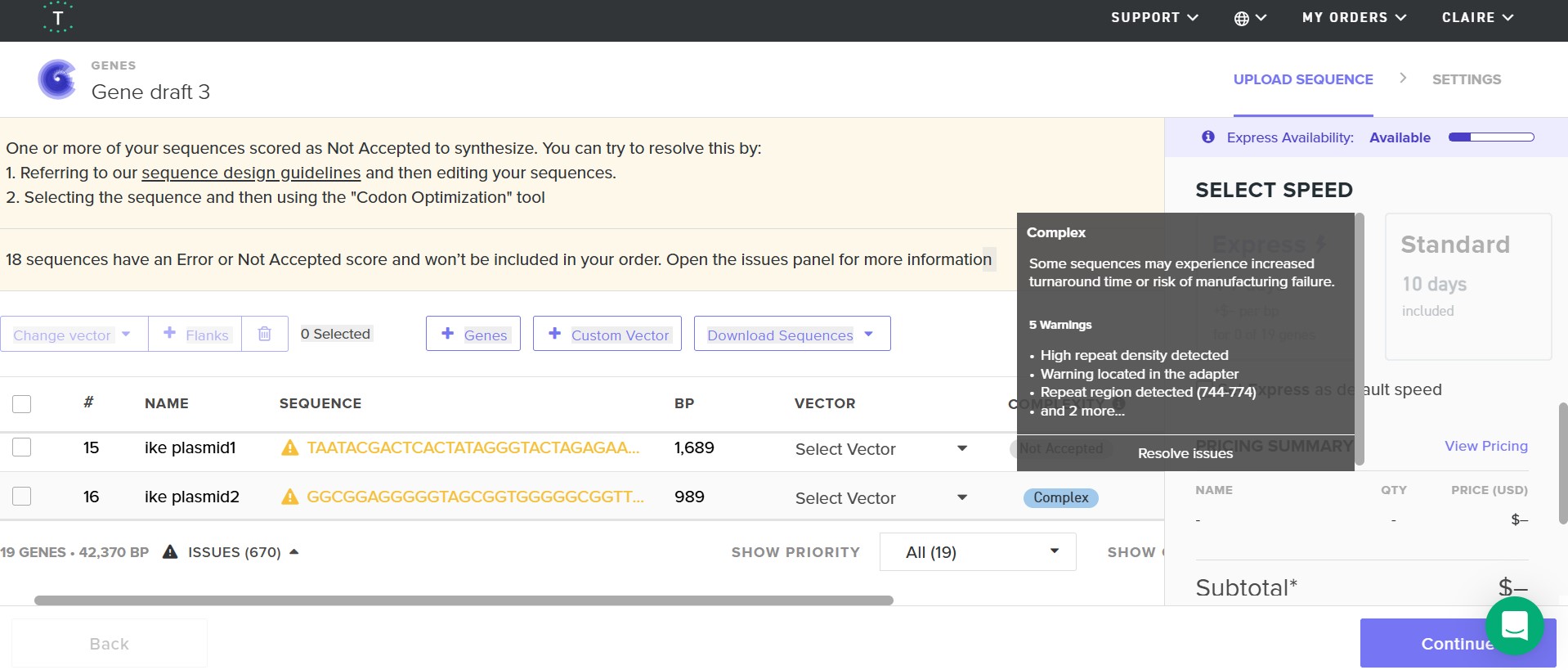
I opted to see if using GenScript a more technically powerful syntethis platform if my construct would be validated, it was. I was warned by GenScript that my construct could face some difficulties due to repitition as expected but I was able to purpose my order. From my research and examples like Snowmax syntethising inaK to use on a big scale in snow canons I know thi construct can be fesable.
GenScript order
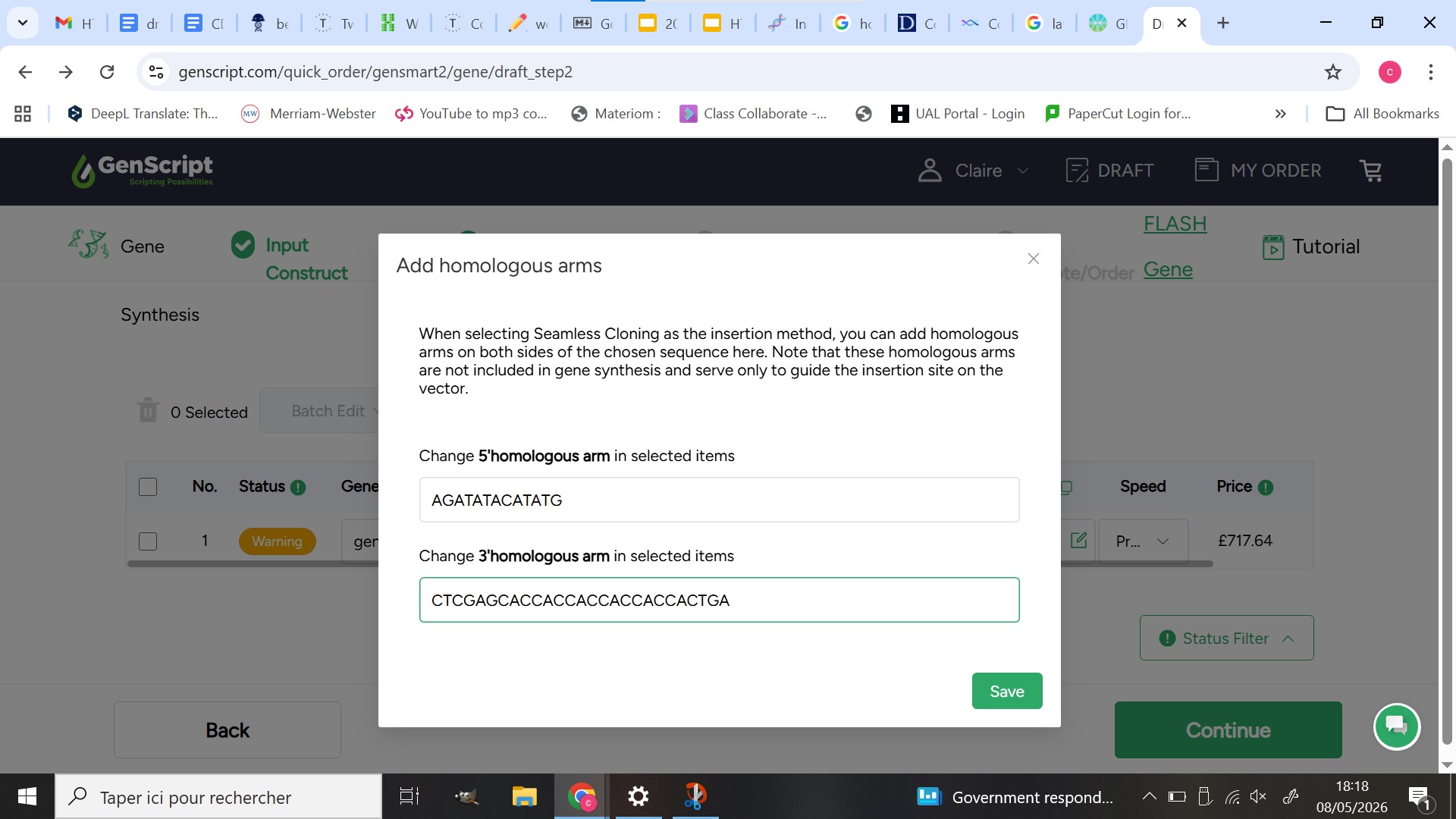
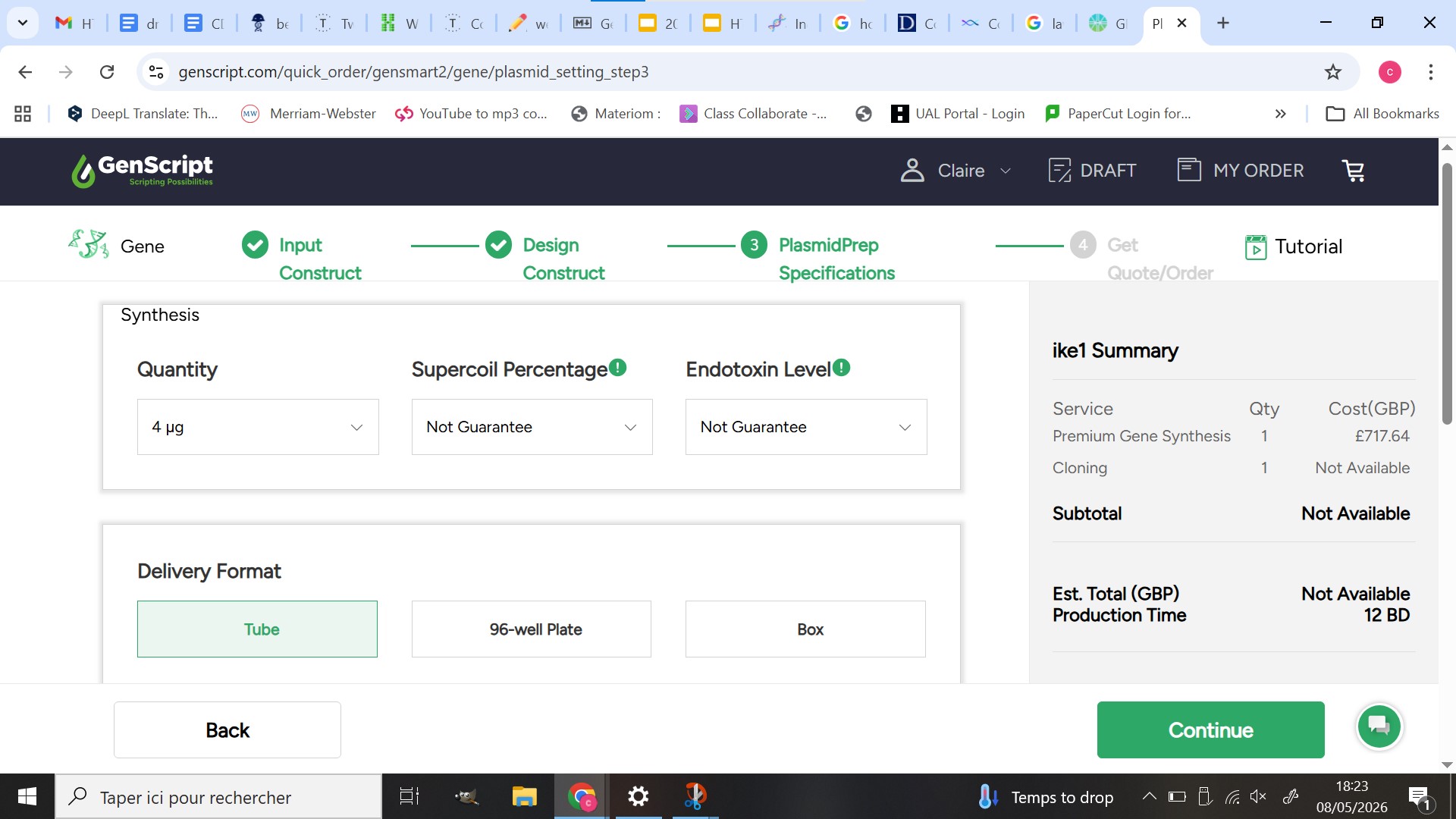
Twist order
As I faced complexity issues with Twist and choose to order my sequence using Genscript I still simulated an order using Twist with a simpler structure using only the sfGFP protein as an example of what a successful Twist order would appear as and to demonstrate that a main domain of my construct can be synthesized using my design.
Final sfGFP construct using pTwist amphigh copy
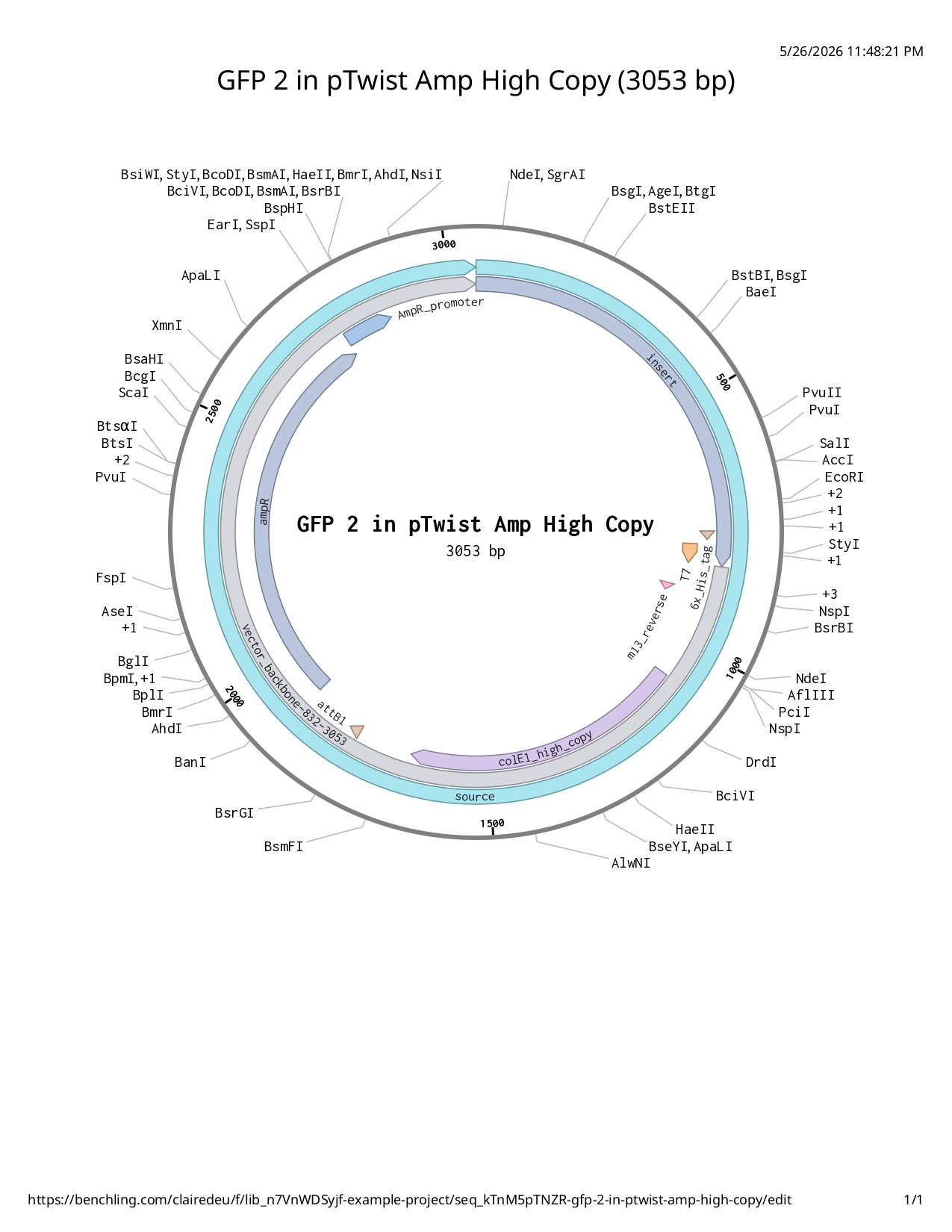
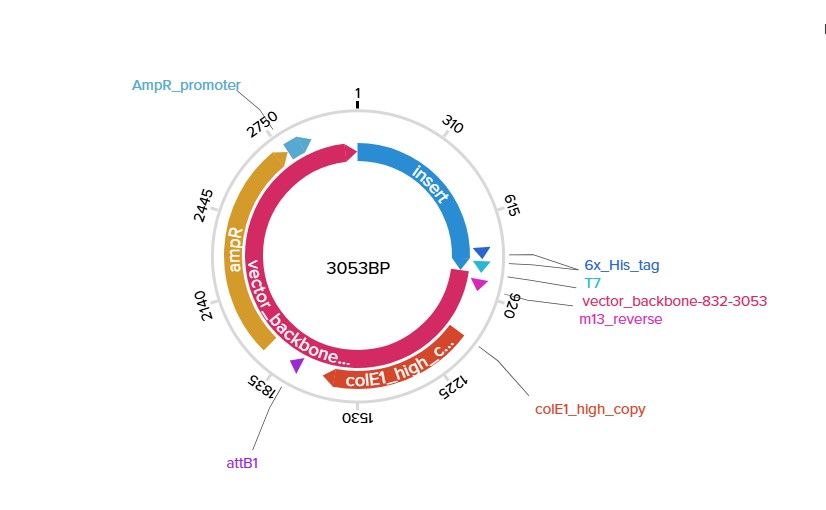
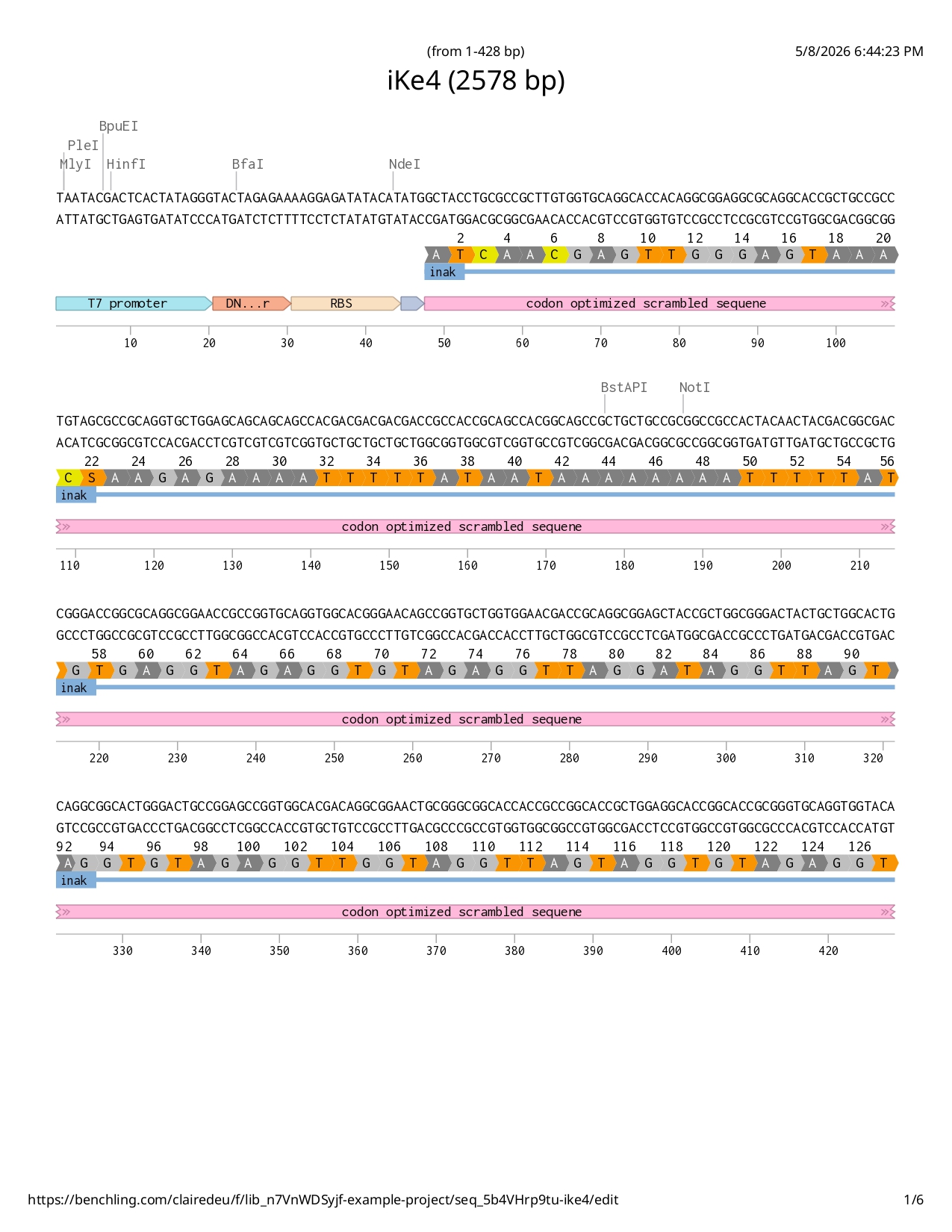
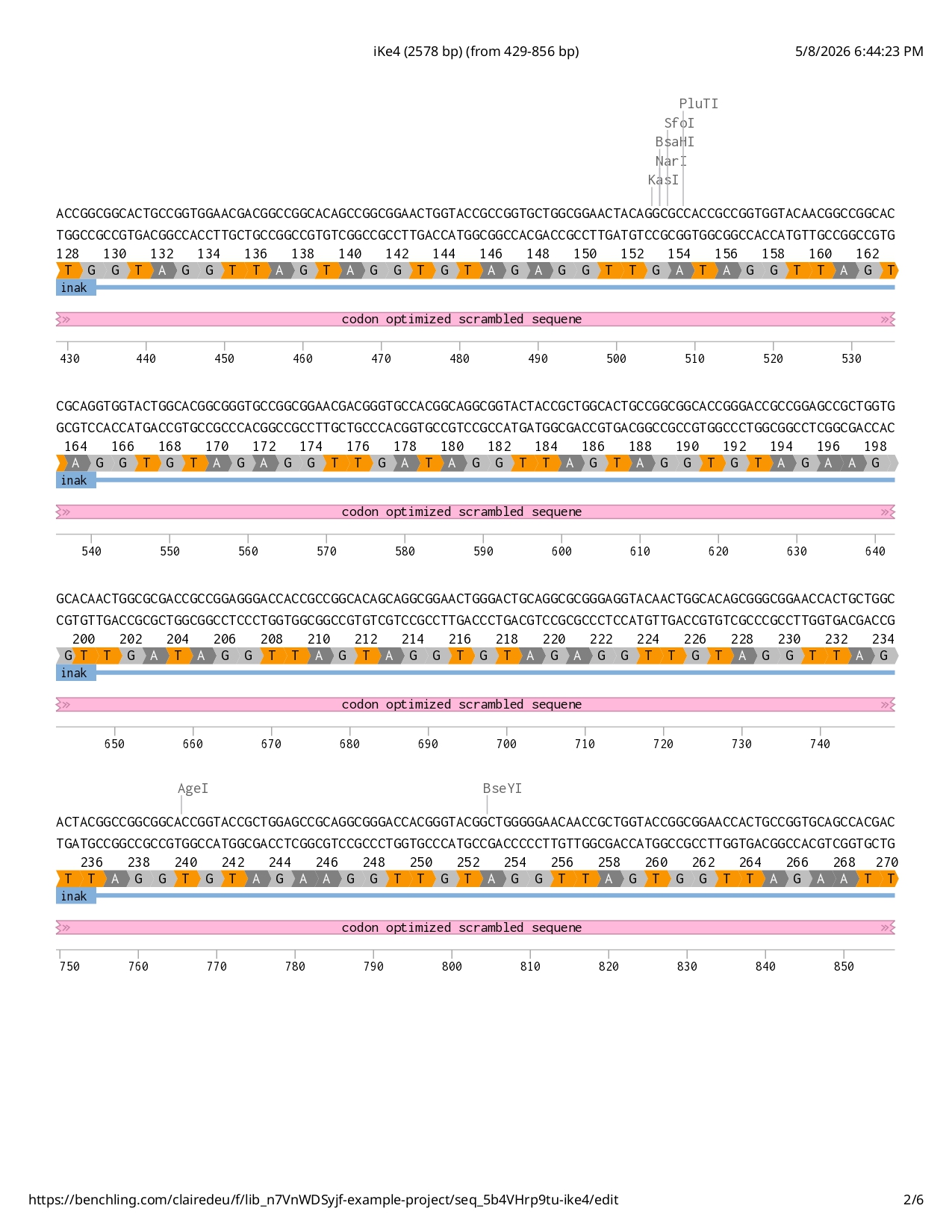
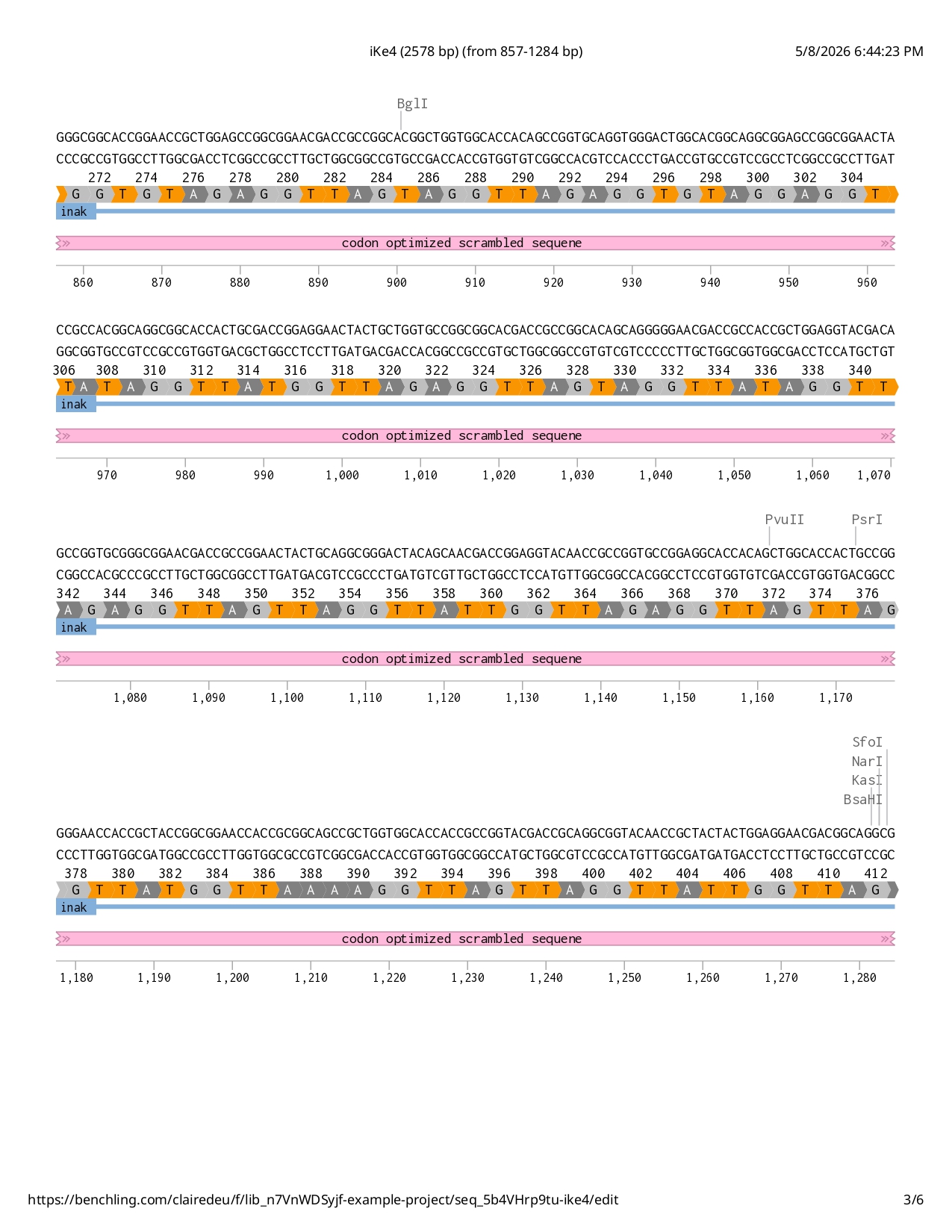
Full iKe construct sequence with translation:
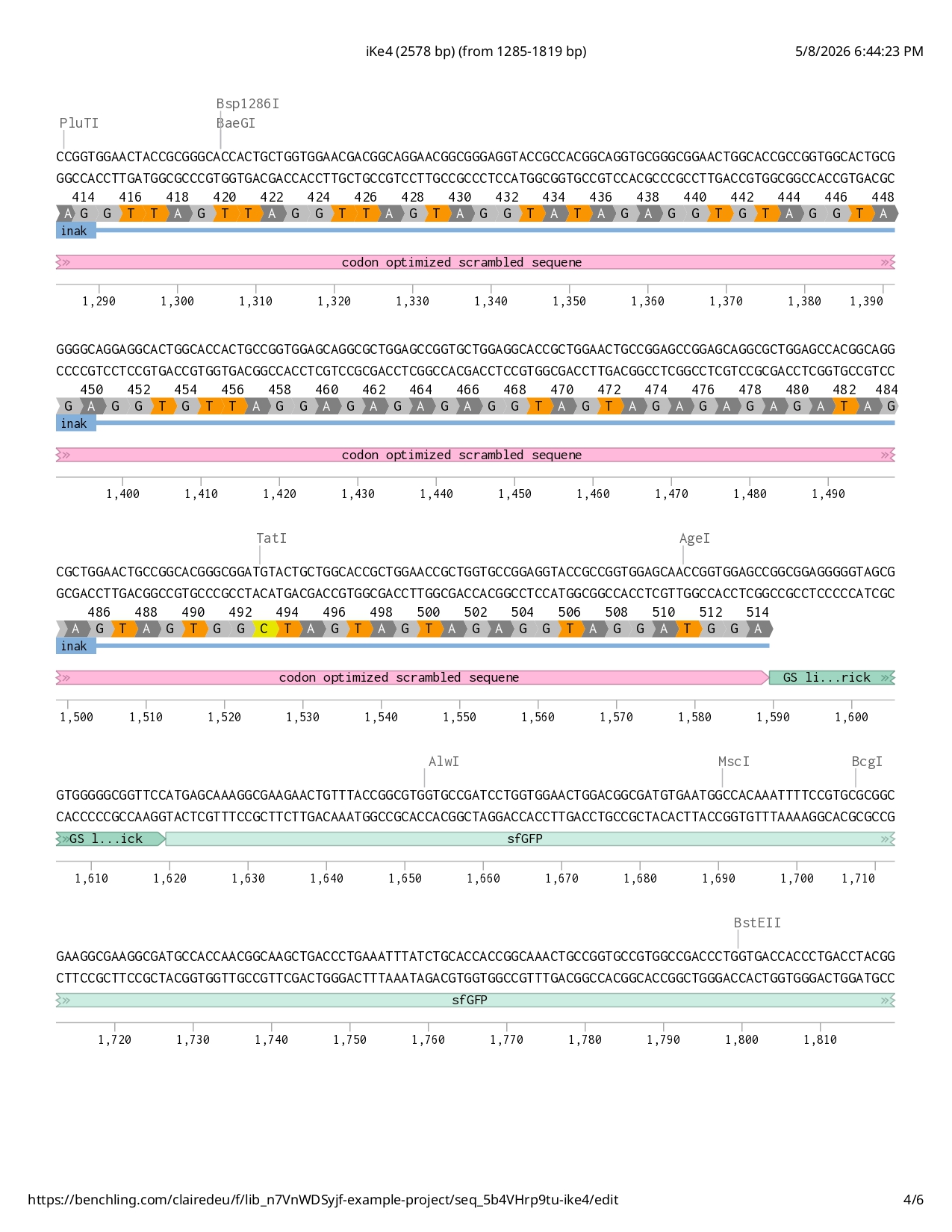
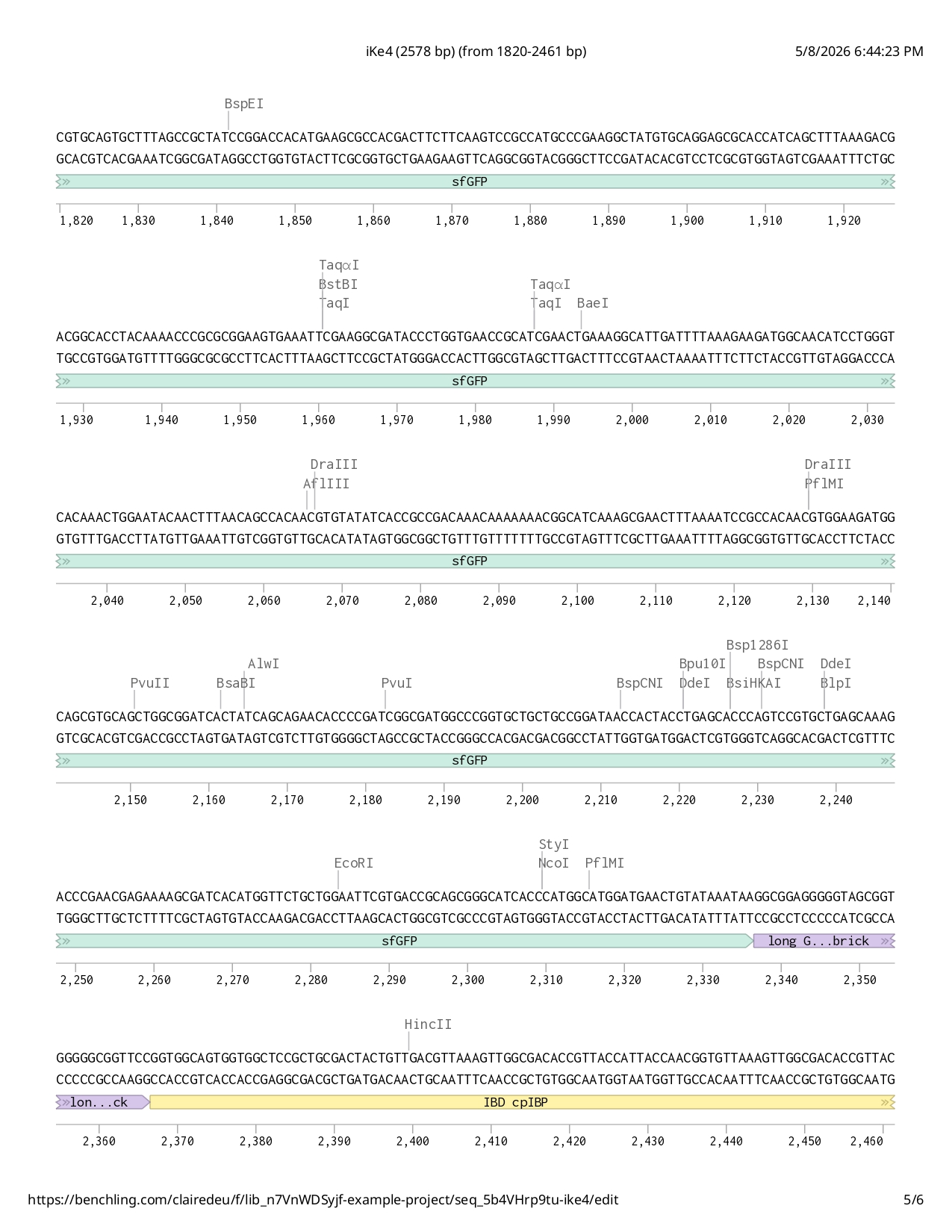
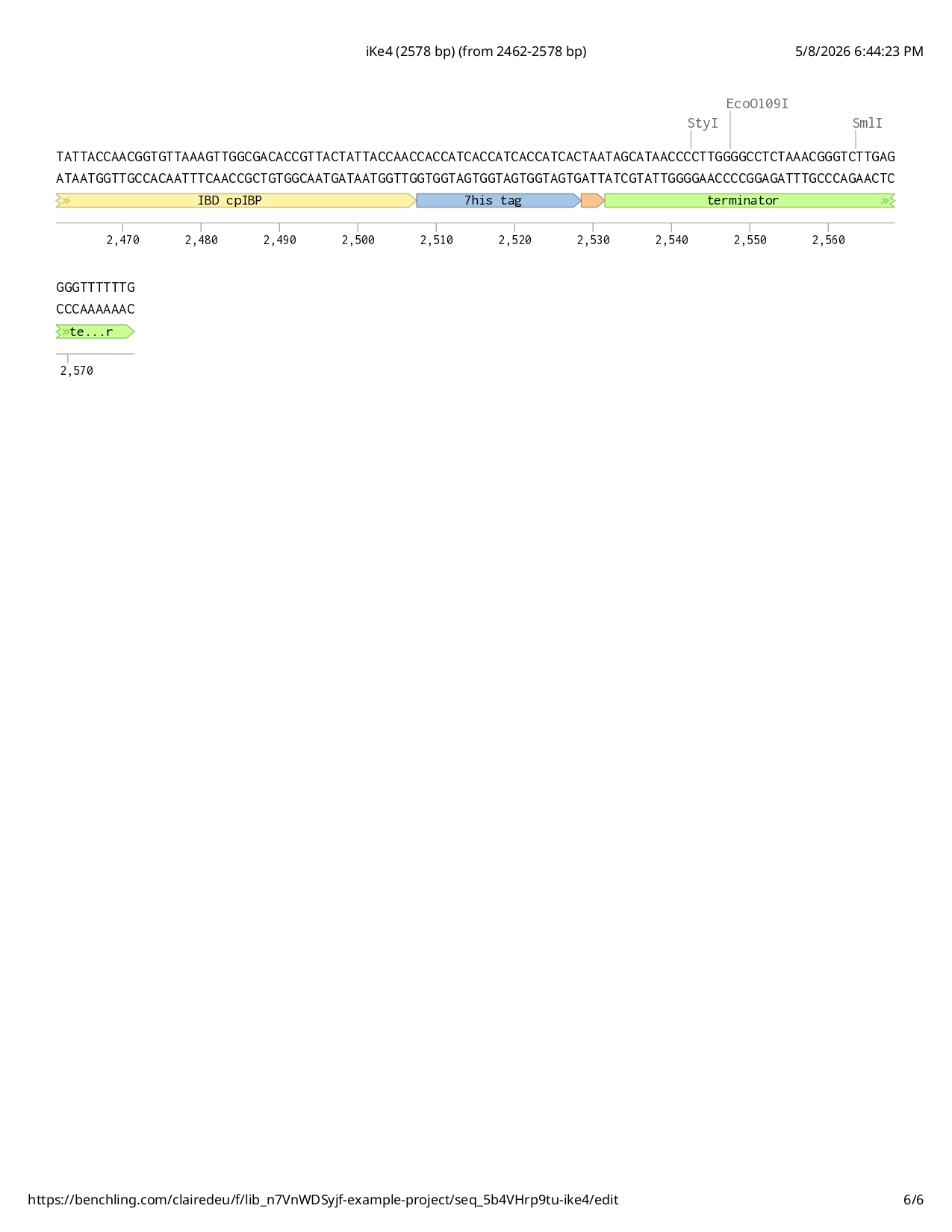
First Design Attempt
Initial project proposal choice

Initial project proposal



Found limitation
It appears a project is already making use of the technology I was planning on innovating on, Snowmax International is already making use of ice nucleating proteins to improve ice catalyzation at higher temperatures in snow cannons to replenish skiing tracks. Therefore, I will keep going with my second aim of modifying certain amino acids of my DNA sequence to improve the function of inaK and find an additional way of pushing further the innovation for this project.
Second Design Attempt & Experimentation Process
Following the limitations found throughout my first design attempts I adjusted my aims accordingly.
Redefining my aims
Aim 1 - modify my inaK sequence to be stronger, more efficient and more stable by combining to the initial design structure an sfGFP tag (contains some UV protective benefits)and an ice binding domain, additionally, add to the master mix a polyol which will help the ice binding domain stick to the surface glacier better.
Aim 2 - increase the scalability and longevity of the project by testing the modified inaK sequence by adding extra Melanin or Scytonemin to increase UV protection and hydrophins to increase better propagation on glaciers and making the ice layer more persistent in time.
According to my aim 1 I designed a protein construct which would boost stability and efficiency of the inaK INP to be used on glaciers.
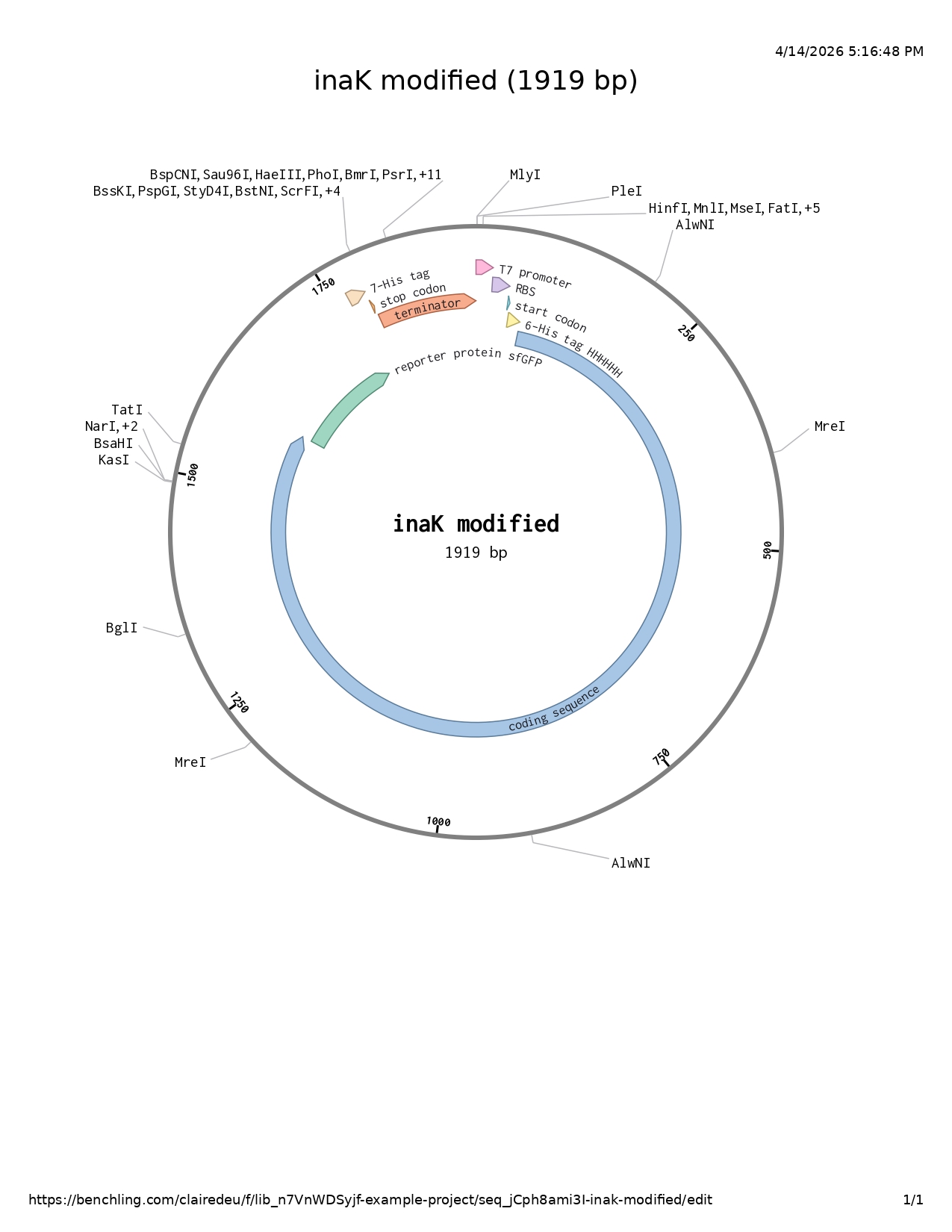
DNA sequencing
I followed the base DNA sequencing structure shown to us in week two and adapted it accordingly to my project.
- T7 promoter
- RBS
- Start codon
- 6His-tag : most common protein purificator
- Codon optimized coding sequence : offers class A nucleation
- Reporter protein sfGFP : increased stability and UV protection with both aspects increase ability to produce ice nucleation and be more resilient to higher temperatures (whether increasing the nucleation point or afterwards resisting warmer external temperatures)
- Glycine serine linker : forms a flexible tether allowing the IBD to move freely and locate the ice surface.
- IBD codon optimized : added to the C-terminal help the INP adhere to the surface of another layer of ice (such as the glacier) preventing INPs to be naturally washed away by meltwater
- CpIBP: compound anchoring the entire complex to the glacier surface
- 7His-tag : protein purification
- Stop codon
- Terminator
Add Sorbitol polyol to inaK master mix which will increase inaK function
Attempt at Twist order
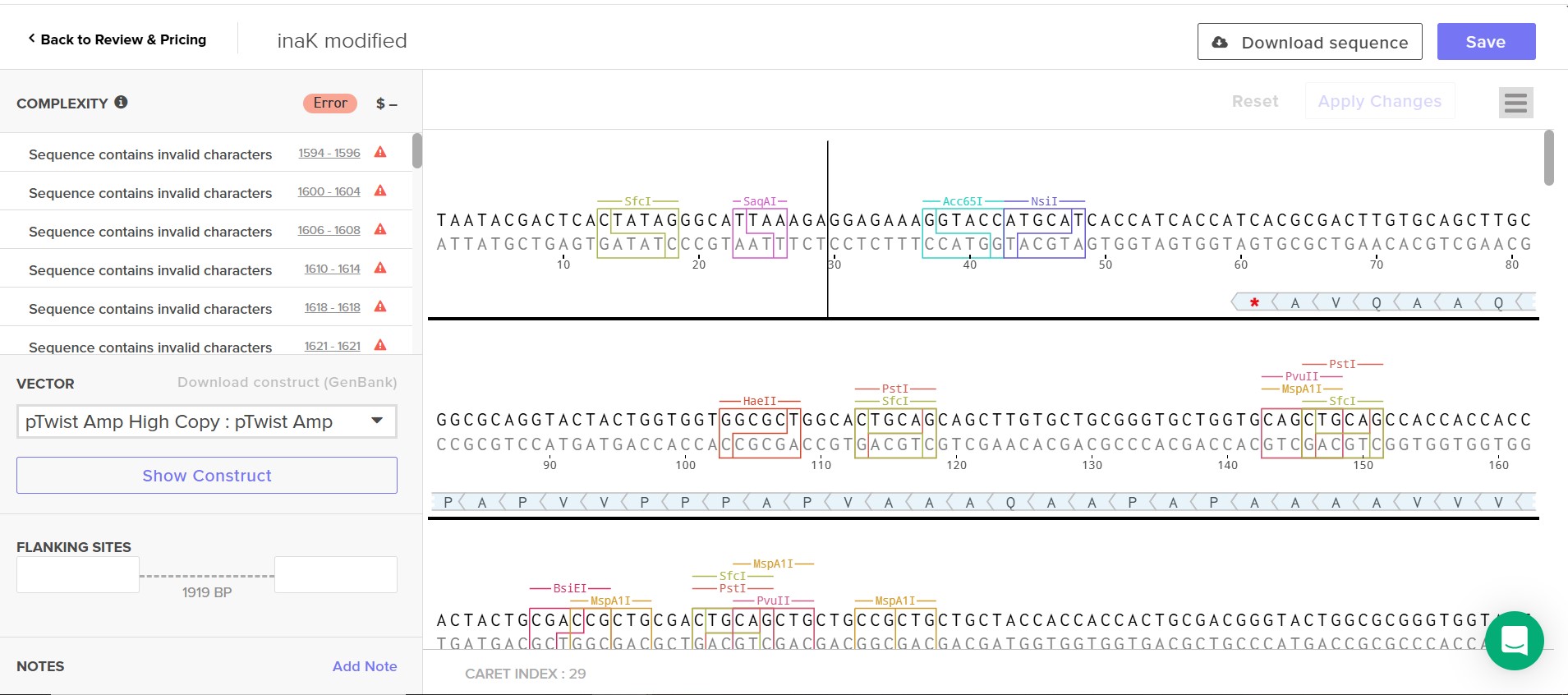
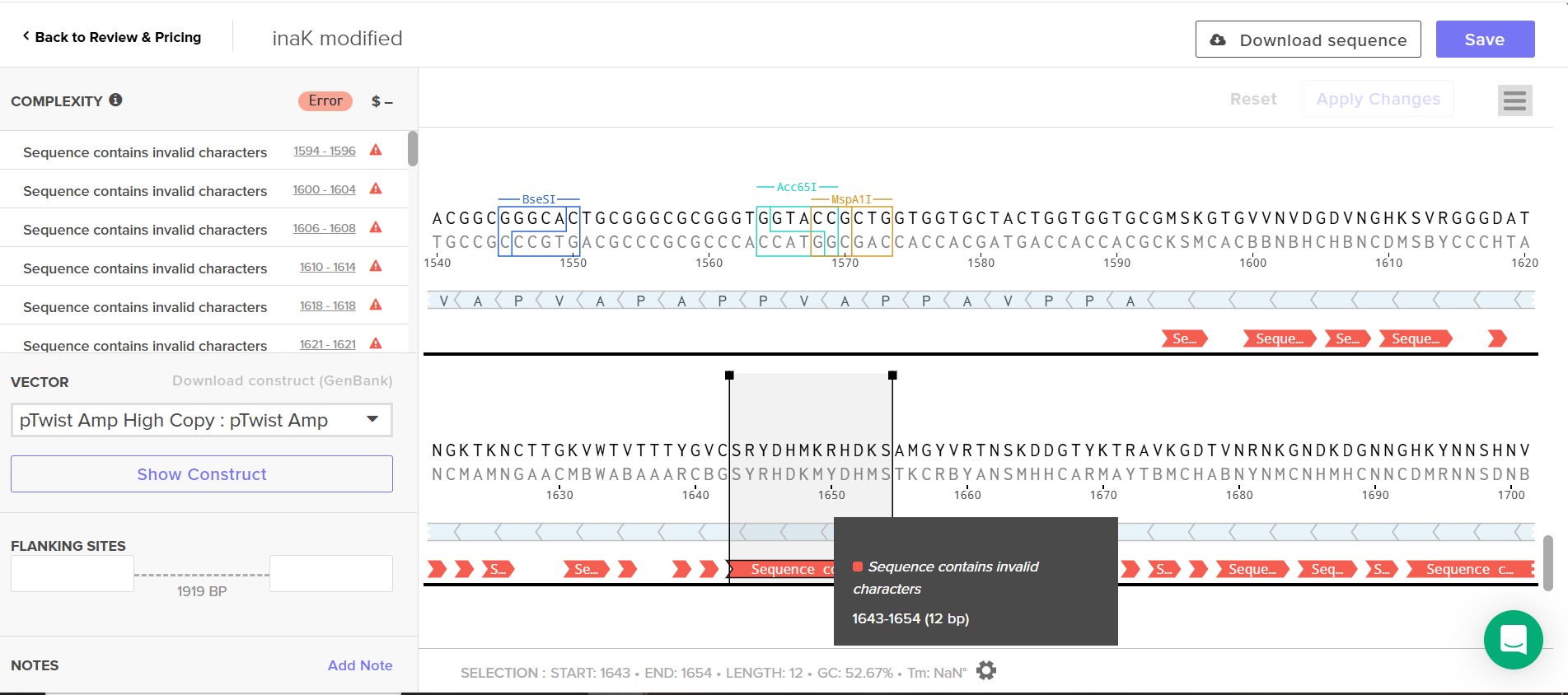
I was consistently faced with Twist issues, at first simple nuclotide sequence errors which I rapidly corrected but then hierarchal design issues where I added linkers to clarify my sequence although and finally I always had the issue of using a repetititive sequence which Twist would not accept its complexity, I did try slpitting my construct in two plasmid simply to see if it would solve the issue but even then it was too complex. I cannot reduce, cut or use only a fragment of my sequence as effective and strong nucleation require a long continuous repetitive motif to organize water molecules and offer a surface for ice crystalization. I attempted to scramble my sequence as much as possible to reduce repitition as much as possible but it still did not work.
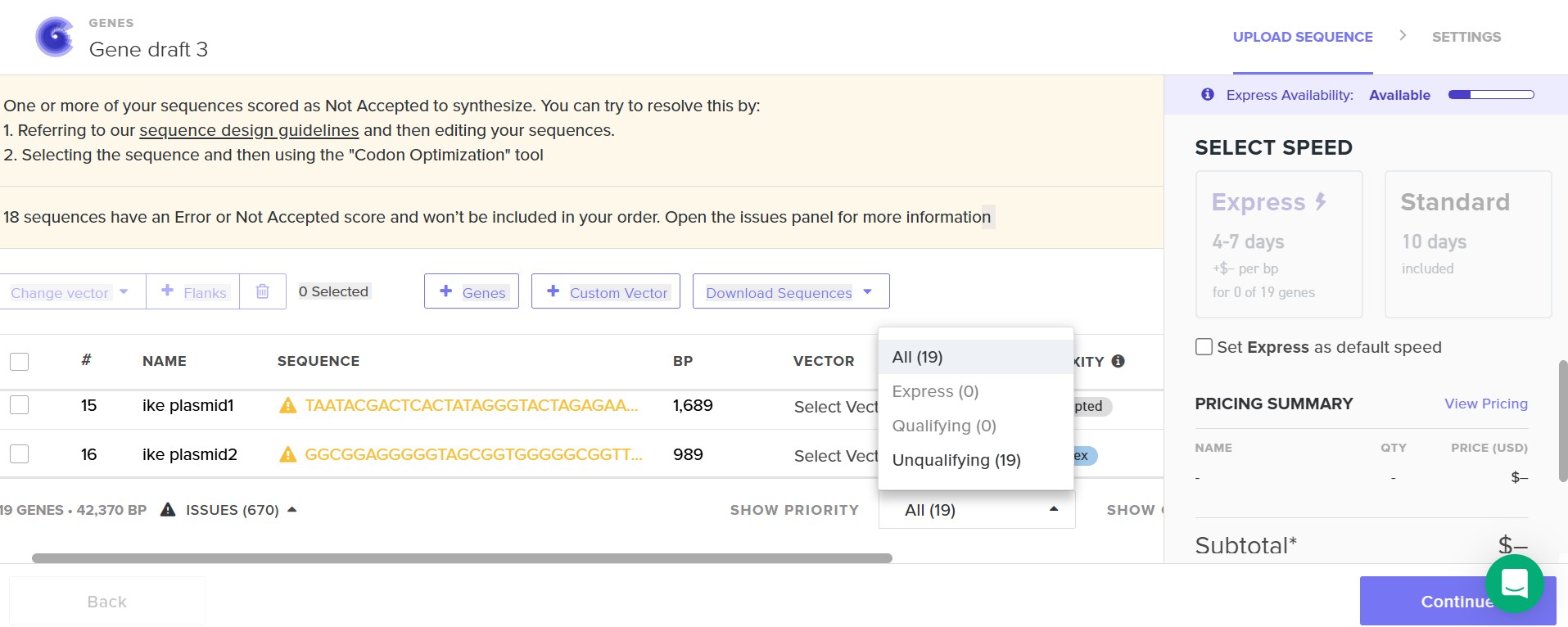
I did 19 construct designs on Benchling and 18 attempts at a Twist order.
From a basic construct simply with my base protein inaK,

to my most promosing construct,

I tried optimizing my construct through Twist, however, it remained to complex
Attempt at seperating construct in two plasmid

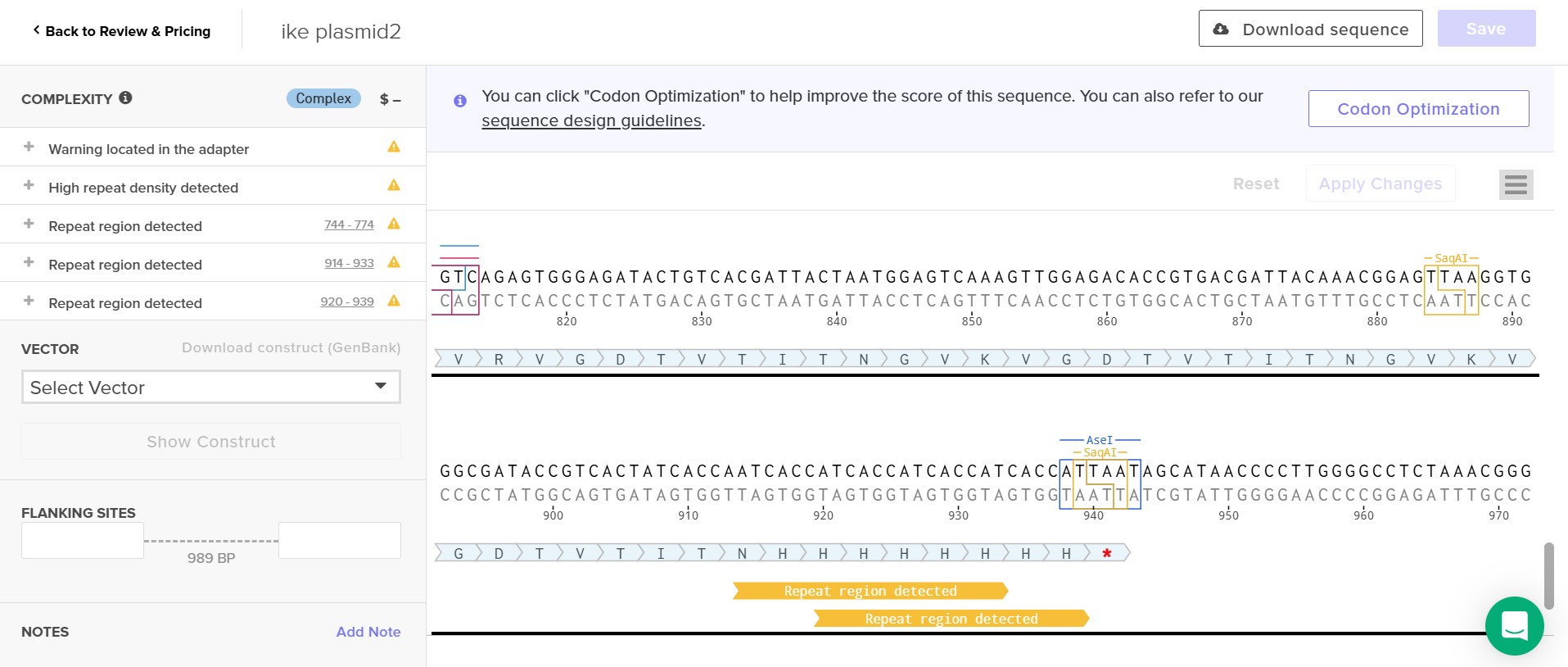
As one can notice the second plasmid is marked as complex but is still not accepted by Twist, the second plasmid still presents too much complexity even as it is much shorter and sipler than the first plasmid
I attempted using a different ice nucleating protein,here the inaQ with less repetitive sections than an inaK. The inaQ sequence on its own was still too complex for Twist to synthesize it.

iKe Design Protocol
Ike Aim 1 experiment and timeline:
Pre, research and development
- In depth research and successful DNA construct design, 4 weeks
- Ordering construct from Twist or Genscript, 10 business days
Phase 1 cloning and growing
- Preparation and transformation : reconstitute ordered plasmid (pET-28a-iKe) and transform into a chemically competent E. coli BL21(DE3) cell via heat shock at 42°C, the pores of the bacteria open and the plasmid can enter and the bacteria have the instructions to produce the protein, 3 days.
- Bacterias are grown on LB-kanamycin plates, only the cells which have accepted the plasmid will survive.
- Grow the successful cells at varying temperatures, single colonies on 5mL LB media, 18°C - 25°C - 37°C to find optimal growth condition and yield, if it is too warm the protein will clump up and if it is too cold it will grow too slowly, 2 days.
Phase 2 making the protein
- Induce IPTG chemical to create a reaction with the T7 promoter to force mass production in iKe protein construct
- Expose construct to fluorescence as a green glow should be seen from the sfGFP, in each cell a bright green halo should be observed as proof of the protein reaching the membrane of the cell. Use a Confocal Laser Scanning Microscope (CLSM), 1 day.
- Cell fractionation, using physical pressure like osmotic shock to break the cells open, centrifuge at high speed to separate the cell content (cytoplasm and periplasm) from the membrane, 3 days.
Phase 3 testing function and efficiency
- Use Western Blot to sort the proteins by size and use antibody to detect 7His-tag and confirm it is found in the outer membrane fraction. Run fractions on a 10% SDS-PAGE gel for Western Blot targeting C-terminal 7His-tag, results should appear at about 85 kDa in the membrane, 2 days.
- Test ice binding by dropping the cells onto a thin layer of ice, the CpIBP should link onto the surface of the ice and prevent ice crystals from growing larger, 2 to 3 days.
- Droplet freezing assay, test the boosted function of the nucleation of inaK by placing droplets of the cells onto a cold plate and progressively lower the temperature, if the inaK nucleates as intended it should force water to freeze at higher sub-zero temperature, 3 days .
- Use a flow cytometer to run cells individually through the laser in order to count precisely how many cells present green fluorescence and provide data on how successful the protein growth was, 1 to 2 days.
Post, quality control ensuring plausibility and reproducibility
- Verify iKe sequence by performing Sanger sequencing on the transformed plasmid using universal T7 primers to confirm the repetitive domain of the inaK has not been affected and recombined in the host.
- Sterility and negative controls will allow to control the difference between the iKe construct ice binding and the background cellular protein behaviour, run an empty vector pET-28a without insert.
- Check proper folding of sfGFP by measuring the excitation spectra (485\nm / 510nm) using a plate reader. A low signal to noise ratio would indicate that the inaK is forcing the sfGFP to misfold into inclusion bodies.
Reference list
Chen, X., Zaro, J.L. and Shen, W.C. (2013) ‘Fusion protein linkers: property, design and functionality’, Advanced Drug Delivery Reviews, 65(10), pp.1357–1369.
Forbes, J., Bissoyi, A., Eickhoff, L., et al. (2022) ‘Water-organizing motif continuity is critical for potent ice nucleation protein activity’, Nature Communications
Gibson, D.G., Young, L., Chuang, R.Y., Venter, J.C., Hutchison, C.A. and Smith, H.O. (2009) ‘Enzymatic assembly of DNA molecules up to several hundred kilobases’, Nature Methods, 6(5), pp.343–345.
Hudait, A., Odendahl, N., Qiu, Y., Paesani, F. and Molinero, V. (2018) ‘Ice nucleation by bacterial proteins: how larger aggregates initiate freezing at warmer temperatures’, Journal of the American Chemical Society, 140(14), pp.4905–4912.
Kim, E.J. and Kim, S. (2010) ‘A versatile microbial cell surface display system using InaK anchor protein’, Methods in Molecular Biology, 605, pp.353–365.
Li, Q., Yan, Q., Chen, J., He, Y., Wang, J., Zhang, H., Yu, Z. and Li, L. (2012) ‘Molecular characterization of an ice nucleation protein from the ice nucleating bacterium Pseudomonas syringae’, Gene, 502(1), pp.1–7.
Pédelacq, J.D., Cabantous, S., Tran, T., Terwilliger, T.C. and Waldo, G.S. (2006) ‘Engineering and characterization of a superfolder green fluorescent protein’, Nature Biotechnology, 24(1), pp.79–88. Schmid, M.A., Jensen, G.J. and Grimm, R. (2010) ‘A new mechanism for bacterial ice nucleation’, Journal of Molecular Biology, 397(3), pp.802–813.
Vance, T.D., Graham, L.A. and Davies, P.L. (2019) ‘Developing a surface-display system for the evaluation of ice-binding proteins’, Journal of Applied Microbiology, 126(3), pp.812–824.
Check List

As a committed listener working in silico I validated the aim for designing a DNA protein construct and tested the order in Twist and Genscript, please refer to project development documentation.
Throughout my final project I made use of a variety of synthetic biology techniques and tools such as DNA sequencing, making a DNA construct using Benchling, genetic databases (NCBI, Uniprot, ENA), , Alpha Fold 3, Pymol, designing a Twist and Genscript order. In order to design my DNA construct I used genetic data bases to source sequences for each element of my construct, I used reverse translation tool, I codon optimized and scrambled my coding sequence, I learn how to build a construct using Benchling where I also annotated each element and exported a circular plasmid, I learned how each piece of the puzzle affects the outcome and success of a construct and guides my choices. Additionally I used Pymol and AlphaFold 3 to visualize and better understand the main domains of my structure. With all the challenges I was faced with using Twist I learn in depth how to use Twist, its limitations and a variety of issues one can be faced with when designing a construct, this really pushed by design and project overall.
Expected results and data
By inputting my research, sequence and protocol into Gemini Assistant I collected mock data and predicted results of my in silico project.
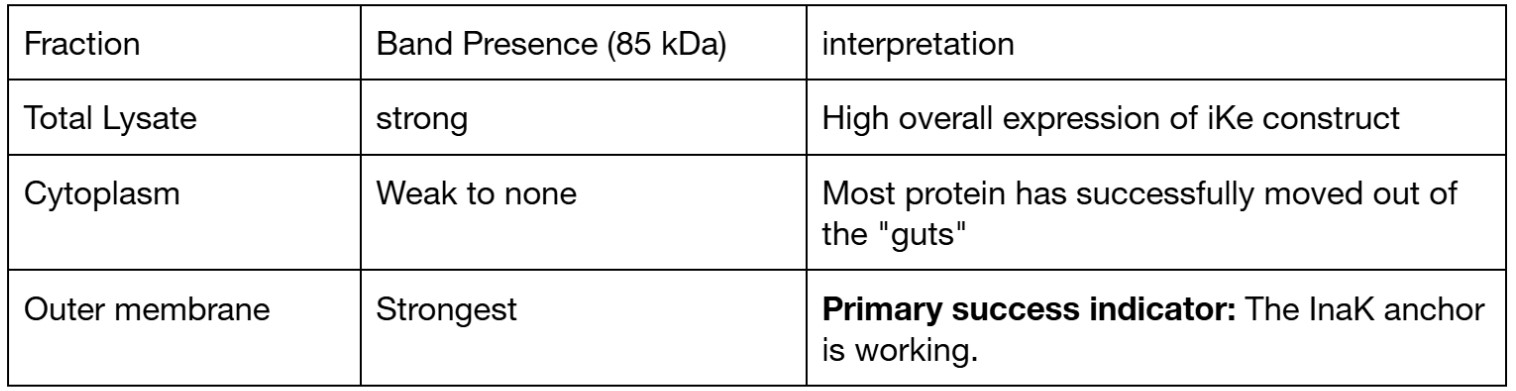
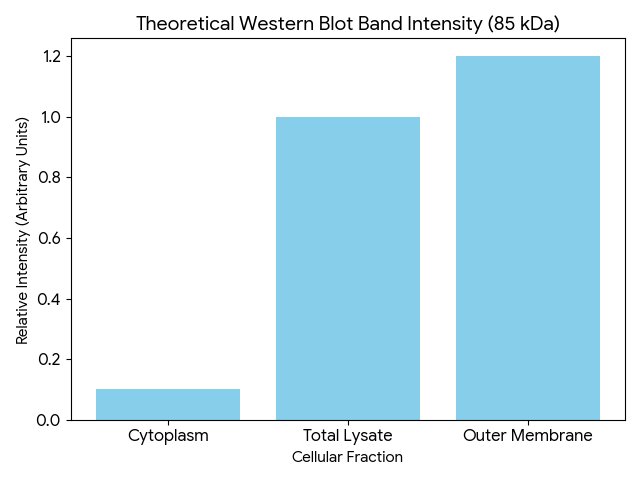
Protein expression and localization Using Western Blot targeting the 7His-tag at the C-terminal I expect to see a clear band at approximately 85 kDa
 Generated by Gemini
Generated by Gemini
 Generated by Gemini
The chart shows a high intensity in the outer membrane area and minimal intensity in the cytoplasm area. This confirms that the inaK is correctly expressing its function in the membrane, as it should for effective ice nucleation.
Generated by Gemini
The chart shows a high intensity in the outer membrane area and minimal intensity in the cytoplasm area. This confirms that the inaK is correctly expressing its function in the membrane, as it should for effective ice nucleation.
Fluorescence tracking Run cells through a flow cytometer to find the correct green fluorescence.
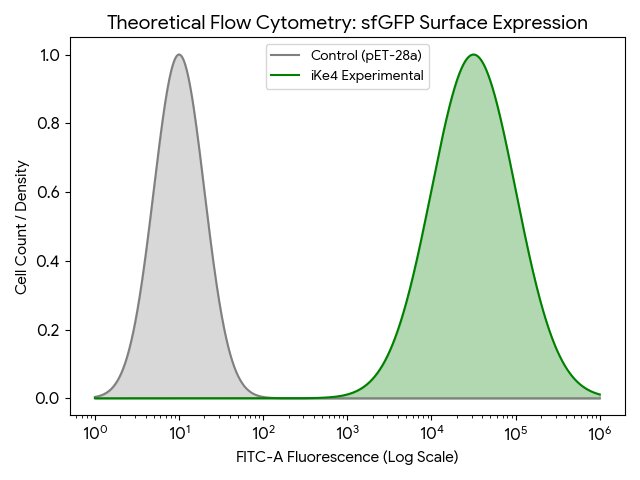 Generated by Gemini
Generated by Gemini
Negative control (pET-28a empty) with a single peak at 10(1). Experimental design with a shift at 10(4) or 10(5). The theoretical result shows >85% of the group should demonstrate high fluorescence confirming the sfGFP domain is folding correctly on the surface of the majority of the cells.
Ice nucleation activity Using the droplet freezing method to test the increased function of the inaK engine. Comparing freezing temperatures of 50 droplets of my culture compared to a control.
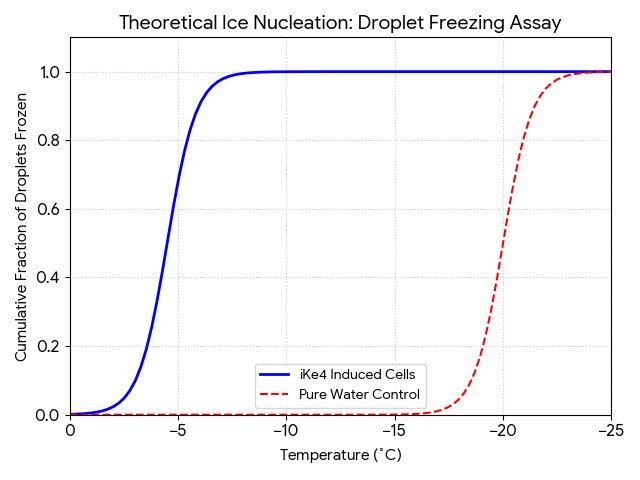 Generated by Gemini
Generated by Gemini
The control in pure water freezes from -18°C to -20°C. The iKe inoculated cells droplets should initiate freezing at -3°C t -5°C. The inaK is creating the necessary ice templates to trigger crystallization on the surface of the cell at high sub zero temperatures.
Ice binding activity Testing the cpIBP domain to measure the size of ice crystals (mean grain size MGS) over time at -6°C
 Generated by Gemini
Generated by Gemini
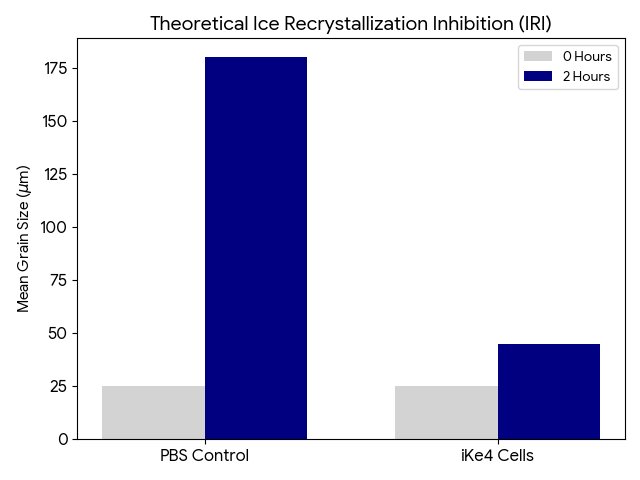 Generated by Gemini
Generated by Gemini
The theoretical data depicts that the iKe inoculated cells should express an MGS reduction of about 75% compared to the control, proving that the cpIBP domain is exposed to the water and successfully linking to the ice crystal borders. The cpIBP is functioning correctly as an anchor between the protein and the ice crystal boundaries.
Theoretically my project appears successful as it presents a positive triple positive logic with successful green fluorescence, water droplets freezing at high sub zero temperatures and a cluster of small ice crystals rather than large plates.
Overall, this data is speculative as my project was run in silico, however, if I were to perform my experiment I would overlay my real data with my predicted results and observe if these align. If my real data matches my theoretical curves then my hypothesis would be supported.
Project cost estimate
As a committed listener working fully in silico I used Gemini by inputting my protocol and project information to create a cost estimate of my project.
Production of my iKe construct
- Genscript order - 717.64£
- Expression Vectors & Bacterial Hosts: Reagents, growth media (LB or TB broth), and competent E. coli or Bacillus subtilis strains for bulk production - 1500£ – 3700£
- Bioreactor / Fermentation Equipment: Access to a pilot-scale bioreactor (10L to 100L) to cultivate high densities of the engineered protein - 11000£ – 30 000£ (or 4 000£ – 7 500£ if outsourced to a contract manufacturing organization).
- Protein Purification Supplies: Centrifuges, lysis buffers, and chromatography columns (FPLC, Ni-NTA resins for His-tagged constructs) to isolate pure iKe proteins from cell debris – 6 000£ – 11 000£
Testing of the construct
- Western Blot (7His-tag detection) via 10% SDS-PAGE to verify expression in the outer membrane fraction (~85 kDa).
- Custom Anti-7His (or Anti-6His) Primary Antibody: To specifically target your C-terminal tag - 250£ – 350£ (per 100 $\mu$L vial).
- Secondary Antibody (HRP-conjugated anti-mouse/rabbit): For chemiluminescent visualization- 100£ – 200£.
- Outer Membrane Fractionation Kit (or Lysozyme + Polyethylene Glycol/Ultracentrifugation reagents): Reagents to selectively isolate the bacterial outer membrane from the cytoplasm and inner membrane - 150£ – 300£.
- SDS-PAGE & Western Blot Consumables: Precast 10% polyacrylamide gels, PVDF or Nitrocellulose membranes, transfer sandwiches, protein ladders (covering 85 kDa explicitly), and ECL chemiluminescence substrate - 200£ – 400£.
- Ice binding & recrystallization inhibitor (IRI) assay
- Specialized Cold Stage or Controlled Temperature Enclosure: To maintain a precise sub-zero microscopic field without ambient room heat melting the thin ice sheet - 1800£ – 5 000£ (to purchase a basic thermal stage) OR 150£ – 400£ instrument rental/core facility access.
- Assay Consumables: High-purity coverslips, cryo-protectants (like sucrose or PBS buffers for background comparison), and a digital camera attachment for an existing microscope to record crystal sizes over 2–3 days - 100£ – 250£.
- Droplet freezing assay
- Automated Micro-Droplet Freezing Plate/Rig: A cooling block capable of precise, linear temperature ramp downs (e.g., down to -20°C at 0.5°C/min) coupled with a video system to catch the exact moment a droplet turns opaque (freezes) - 2 000£ – 6 000£ to buy/build, or 400£ in custom parts using a thermoelectric Peltier cooling module if built DIY.
- Assay Surface Modifiers: Hydrophobic glass slides or silicone oil matrix to prevent the droplets from prematurely nucleating on the plate surface itself rather than via the InaK engine - 80£ – 150£.
- Flow cytometry quantification
- Flow Cytometer Core Facility Fees: Standard charge for using a shared university analyzer machine (e.g., BD FACSCanto or similar), typically billed hourly - 60£ – 100£ per hour (Estimated 4 hours total for replicates: 250£ – 450£).
- Cytometry Consumables: 96-well round-bottom plates or specialized polystyrene tubes, calibration beads, and cell-staining/dilution buffers - 100£ – 200£.
Quality control
- Sanger Sequencing (with Universal T7 Primers): Forward and reverse reads to verify that the unstable, repetitive domains of InaK haven’t recombined out of your plasmid sequence - 5£ – 10£ per reaction (Assuming 5 clone validations = 40£ – 60£).
- Empty Vector Control Culture: Reagents to maintain and express the control group containing the blank pET-28a plasmid (kanamycin selection antibiotics, cell growth media) - 40£ – 80£.
- Fluorescence Microplate Reader Access/Consumables: Black-walled 96-well plates (essential to prevent signal bleed-through when measuring 485nm excitation / 510nm emission spectrums) and instrument time - 100£ – 200£.
The price would also vary if working with university labs or public institution labs.
Bibliography
Anacona, P.I., Kinney, J., Schaefer, M., Harrison, S., Wilson, R., Segovia, A., Mazzorana, B., Guerra, F., Farías, D., Reynolds, J.M. and Glasser, N.F. (2018). Glacier protection laws: Potential conflicts in managing glacial hazards and adapting to climate change. Ambio, 47(8), pp.835–845. doi:https://doi.org/10.1007/s13280-018-1043-x.
Arnold, D.L. and Preston, G.M. (2019). Pseudomonas syringae: enterprising epiphyte and stealthy parasite. Microbiology, 165(3), pp.251–253. doi:https://doi.org/10.1099/mic.0.000715.
Boztas, S. (2024). Pumped up: will a Dutch startup’s plan to restore Arctic sea-ice work? The Guardian. [online] 27 Feb. Available at: https://www.theguardian.com/environment/2024/feb/27/climate-crisis-arctic-ecosystems-environment-startup-plan-pump-restore-melting-sea-ice-caps.
Biotreks (2021) ‘Ice nucleation proteins – a synthetic pathway to alleviate ice loss’, Biotreks, (e202111).
Christner, B.C., Morris, C.E., Foreman, C.M., Cai, R. and Sands, D.C. (2008). Ubiquity of Biological Ice Nucleators in Snowfall. Science, 319(5867), pp.1214–1214. doi:https://doi.org/10.1126/science.1149757.
Davies, P.L. (2014) ‘Ice-binding proteins: a remarkable capacity to adapt for life at cold temperatures’, Biochemical Journal, 458(1), pp. 9–20. doi: 10.1042/BJ20131291.
Dr. Tobias Weidner, Dr. Janine Fröhlich-Nowoisky (2016). The effect of bacterial ice nuclei. [online] Www.mpg.de. Available at: https://www.mpg.de/10470442/ice-formation-bacteria-syringae.
Experimental Data Figure (2026) Intact Mass Spectra: Native vs. Denatured sfGFP Analysis.
Experimental Data Figure (2026) Sequence Coverage Report: 88% Confirmation of sfGFP.
Experimental Data Figure (2026) CDMS Spectrum of KLH Oligomeric States.
Forbes, J., Bissoyi, A., Eickhoff, L., et al. (2022) ‘Water-organizing motif continuity is critical for potent ice nucleation protein activity’, Nature Communications
Garnham, C.P. et al. (2011) ‘A conserved water-organizing motif in ice-nucleating proteins’, Molecular Microbiology, 79(6), pp. 1419–1427. doi: 10.1111/j.1365-2958.2011.07546.x.
Genevaenvironmentnetwork.org. (2024). Unprecedented Rates of Mountain Glacier Melting | Glaciers and the Role of Geneva. [online] Available at: https://www.genevaenvironmentnetwork.org/fr/ressources/nouvelles/unprecedented-rates-of-mountain-glacier-melting/.
GoldBio (n.d.) The Extraordinary Bacterial Proteins That Make Snow. Available at: https://www.goldbio.com/blogs/articles/the-extraordinary-bacterial-proteins-that-make-snow (Accessed: 25 April 2026).
Govindarajan, A.G. and Lindow, S.E. (1988) ‘Size of bacterial ice-nucleation sites measured in situ by radiation inactivation’, Proceedings of the National Academy of Sciences (PNAS), 85(5), pp. 1334–1338.
Hudait, A. et al. (2024) ‘Hierarchical assembly and environmental enhancement of bacterial ice nucleators’, Proceedings of the National Academy of Sciences (PNAS), 121(18), p. E2409283121.
ISPI. (2024). Mr. Matterhorn and Its Glaciers – A Critical Exploration of the Protection of Glaciers through Rights of Nature | ISPI. [online] Available at: https://www.ispionline.it/en/publication/mr-matterhorn-and-its-glaciers-a-critical-exploration-of-the-protection-of-glaciers-through-rights-of-nature-187115 [Accessed 13 May 2026].
James Dalton, Global Head, Water and Wetlands Team, IUCN (2025). Protecting glaciers – our most effective natural water manager. [online] IUCN. Available at: https://iucn.org/blog/202503/protecting-glaciers-our-most-effective-natural-water-manager.
Jung, H.C. et al. (1998) ‘Expression of Candida antarctica lipase B on the surface of Escherichia coli using InaK anchoring motif’, Enzyme and Microbial Technology, 22(5), pp. 348–354.
Lee, C.Y., et al. (2015) ‘Controlled ice nucleation by ice-nucleating proteins for the cryopreservation of complex biological systems’, Biomaterials, 38, pp. 11–21. doi: 10.1016/j.biomaterials.2014.10.050.
Li, Q. et al. (2009) ‘Surface display of Vitreoscilla hemoglobin on Escherichia coli using InaK-N and its effects on cell growth’, Letters in Applied Microbiology, 49(1), pp. 71–76.
Ling, M. L. et al. (2018) ‘The constructive role of protein repeats in ice nucleation’, Nature Communications, 9, p. 3314.
Lukas, M. et al. (2025) ‘A New Class of Fungal Ice-Nucleating Proteins with Bacterial Ancestry’, ChemRxiv. doi: 10.26434/chemrxiv-2025-73058.
Lindow, S.E. et al. (1989) ‘Relationship between Ice Nucleation Frequency and inaZ Protein Content in Escherichia coli’, Molecular Plant-Microbe Interactions, 2(5), pp. 262–272.
McDonough, F. (n.d.). What is Cloud Seeding? [online] Desert Research Institute. Available at: https://www.dri.edu/cloud-seeding-program/what-is-cloud-seeding/.
NANGERONI, G. and VANNI, M. (1963). THE ACTIVITIES OF THE ITALIAN GLACIOLOGICAL COMMITTEE ON THE OCCASION OF THE INTERNATIONAL GEOPHYSICAL YEAR (IGY) / L’activité du Comité Glaciologique Italien à l’occasion de l’Année Géophysique Internationale (A. G. I.). International Association of Scientific Hydrology. Bulletin, 8(3), pp.97–101. doi:https://doi.org/10.1080/02626666309493342.
O’Sullivan, D. et al. (2016) ‘The influence of pH, ionic strength and soluble organics on the ice nucleating ability of Pseudomonas syringae’, Atmospheric Chemistry and Physics, 16(11), pp. 7443–7454
Pandey, R. et al. (2016) ‘Ice-nucleating bacteria control the order and dynamics of interfacial water’, Science Advances, 2(4), p. e1501630. doi: 10.1126/sciadv.1501630.
Pédelacq, J.D., Cabantous, S., Tran, T., Terwilliger, T.C. and Waldo, G.S., 2006. Engineering and characterization of a superfolder green fluorescent protein. Nature Biotechnology, 24(1), pp.79-88. (Note: This reference validates the use of sfGFP for your visual tracing and quality control folding assay).
Ponte di Legno Tonale. (n.d.). The protection of Presena Glacier. [online] Available at: https://www.pontedilegnotonale.com/en/pontedilegno-tonale-what-to-see/the-protection-of-presena-glacier/.
Rgs.org. (2026). RGS Explore Grants. [online] Available at: https://www.rgs.org/exploration/grants/expedition-grants/rgs-explore-grants [Accessed 12 May 2026].
Roeters, S. J. et al. (2024) ‘Polyol-Induced 100-Fold Enhancement of Bacterial Ice Nucleation Efficiency’, The Journal of Physical Chemistry C, 128(15).
Roeters, S.J., Golbek, T.W., Bregnhøj, M., Drace, T., Alamdari, S., Roseboom, W., Kramer, G., Šantl-Temkiv, T., Finster, K., Pfaendtner, J., Woutersen, S., Boesen, T. and Weidner, T. (2021). Ice-nucleating proteins are activated by low temperatures to control the structure of interfacial water. Nature Communications, [online] 12(1), p.1183. doi:https://doi.org/10.1038/s41467-021-21349-3.
Schmid, D. (2026) Glacier Blankets Could Help Prevent Melting. [Online Video]. 25 April. Available at: https://www.youtube.com/watch?v=hKT_SGK2qtY (Accessed: 27 April 2026).
Schmid, J., Sperl, J.M. and Sieber, V., 2015. Cell-free protein synthesis: an emerging technology for the production of tailored proteins and biopolymers. Applied Microbiology and Biotechnology, 99(21), pp.8893-8907.
Schoborg, J.A. et al. (2014) ‘Aqueous two-phase system (ATPS) for direct fractionation of proteins from cell-free protein synthesis’, Biotechnology and Bioengineering, 111(12), pp. 2405–2415.
Silverman, A.D. et al. (2020) ‘Cell-free gene expression: an expanded repertoire of applications’, Nature Reviews Genetics, 21(3), pp. 151–170. doi: 10.1038/s41576-019-0186-3.
Shi, H. et al. (2015) ‘A novel surface display system using the ice nucleation protein InaK-N as an anchor for the directed evolution of a highly active organophosphorus hydrolase’, Applied and Environmental Microbiology, 81(15), pp. 5128–5135.
Snomax.com. (2015). FAQ - Snomax. [online] Available at: https://www.snomax.com/faq.html. Snomax International (2026) The Science of Snomax: Maximizing Snow Production Efficiency. [Online Technical Bulletin].
Steroplast Healthcare (2022). How Do Instant Ice Packs Work? [online] www.steroplast.co.uk. Available at: https://www.steroplast.co.uk/knowledge-base/how-do-ice-packs-work.html.
Turner, M.A., Arellano, F. and Kozloff, L.M., 1991. Components of ice nucleation structures of Pseudomonas syringae, Erwinia herbicola, and Escherichia coli in plasma membranes. Journal of Bacteriology, 173(21), pp.6874-6880. (Note: This supports your Phase 3 protocol verifying the fractionated outer-membrane localization of the InaK engine).
UN (2021). UN Decade on Restoration. [online] UN Decade on Restoration. Available at: https://www.decadeonrestoration.org/. www.cnr.it. (n.d.). Home | Consiglio Nazionale delle Ricerche. [online] Available at: https://www.cnr.it/en.
Un-glaciers.org. (2026). Glacier Preservation is the Key to Ensuring the Security of Water, Energy, and Environmental Resources. [online] Available at: https://www.un-glaciers.org/en/articles/glacier-preservation-key-ensuring-security-water-energy-and-environmental-resources. USNSJ (n.d.) ‘Wonders of the Invisible World: Pseudomonas syringae, the Ice Maker’, University of Southern North Science Journal, 2(2).
Wadokar, T. (2025). Snow Seeds - Tanay Wadodkar - UAL Showcase. [online] Arts.ac.uk. Available at: https://ualshowcase.arts.ac.uk/project/635735/cover [Accessed 9 Feb. 2026].
Walser, A. et al. (2024) ‘Fungal ice nucleation proteins open new pathways for weather modification and biopreservation’, Science Advances, 10(12), p. eadl1234. doi: 10.1126/sciadv.adl1234.
Warren, G.J. and Wolber, P.K., 1991. Molecular biology of microbial ice nucleation. Biomedical and Life Sciences, 55(3), pp.1043-1048.
Wellpott, V. and Wellpott, V. (2025). Glacier protection with geotextiles – A sustainable solution for the future. [online] Naue - Geosynthetics | Digtal Engineering Software | Installation services. Available at: https://www.naue.com/glacier-protection-with-geotextiles-a-sustainable-solution-for-the-future/.
Zachariassen, K.E. and Kristiansen, E., 2000. Ice-nucleating agents: definitions, methods of structural assay and mechanisms of action. Cryobiology, 41(4), pp.257-279. (Note: This research provides the methodological framework for your droplet freezing and Ice Recrystallization Inhibition assays).
