Projects
Final projects:
- A remote-executable HTGAA final project: design three Twist constructs and test a cell-free split-GFP biosensor for the PARP1-HPF1 protein interaction.
Student: Constantin Convalexius
Node: Lifefabs Institute
Course: HTGAA 2026
Project type: DNA construct design + cell-free expression + protein-protein interaction biosensor
Wet-lab scope: 3 Twist clonal genes + 1 Ginkgo Cloud Lab cell-free expression assay
I am building a cell-free split-GFP biosensor to test whether a PARP1 catalytic-domain construct and its partner HPF1 can be co-expressed in an E. coli cell-free system and generate green fluorescence when the two proteins interact.
This project is not a full rejuvenation experiment. It does not directly measure cellular reprogramming, epigenetic age, PARP1 catalytic activity, or gene-regulatory changes in living cells.
The realistic experiment I can run through HTGAA, Twist, and Ginkgo Cloud Lab is narrower and cleaner:
Can I design and build a three-construct split-GFP biosensor that reports the PARP1-HPF1 interaction in a cell-free reaction?
That is still valuable. Before testing a large biological hypothesis in mammalian cells, I first need a working molecular tool. This project builds that tool.
Partial cellular reprogramming can reverse some molecular features of aging, but the mechanisms that separate rejuvenation from loss of cell identity remain incompletely understood. Recent work by Yücel et al. identified conserved master regulators associated with reprogramming-induced rejuvenation, including EZH2 and PARP1. One striking observation is that the catalytically dead EZH2-Y726D mutant can still support rejuvenation-associated effects, suggesting that some regulators may act through non-canonical structural or scaffolding roles rather than only through enzyme activity.
My final project builds a practical experimental tool to begin studying that idea in a remote, HTGAA-compatible format. Instead of attempting a full mammalian reprogramming experiment, which would require cell culture, sequencing, and a much larger budget, I focus on one molecular interaction: PARP1 and HPF1. HPF1 is a known binding partner of the PARP1 catalytic domain and helps direct PARP1-dependent ADP-ribosylation biology. I designed three Twist clonal gene constructs: PARP1 catalytic domain wild type fused to GFP11, PARP1 catalytic domain E988K mutant fused to GFP11, and full-length HPF1 fused to GFP1-10. These constructs are designed for E. coli cell-free protein synthesis at Ginkgo Cloud Lab. Computational validation of the construct design was performed using AlphaFold 3 prediction of the binary PARP1cat-GFP11 / HPF1-GFP1-10 complex, which confirmed (ipTM = 0.71, pTM = 0.64) that the designed fusion proteins recapitulate the known PARP1-HPF1 binding mode and that the split-GFP halves can be brought into reassembly geometry.
The broad objective is to create a working cell-free split-GFP biosensor for the PARP1-HPF1 interaction. My hypothesis is that co-expression of HPF1-GFP1-10 with PARP1cat-GFP11 will produce fluorescence above background if the proteins bind and bring the split-GFP fragments together. The expected outcome is not a direct reprogramming result, but a validated construct-and-assay pipeline that can be expanded later to more regulators and more rigorous functional assays.
The biological motivation comes from rejuvenation and reprogramming literature, but the HTGAA contribution is the engineering pipeline:
This is the HTGAA part: taking a biological idea and turning it into a buildable synthetic biology experiment.

Ginkgo Nebula. The wet-lab part of this project is designed for Ginkgo’s automated cloud lab infrastructure. Instead of manually pipetting every reaction myself, the DNA constructs can be tested through robotic liquid handling, cell-free expression reactions, and plate-reader measurement inside this kind of automated experimental platform.
Innovation. This project operationalizes the catalytic-versus-scaffolding distinction, motivated by the catalytically-dead EZH2-Y726D rejuvenation phenotype, with a buildable, remotely-executable wet-lab assay rather than a hypothesis on paper. It adapts the tripartite split-GFP system of Cabantous and Waldo (2005) from mammalian PPI work into a cell-free format compatible with Ginkgo Cloud Lab, and produces an open construct set that can be extended to the remaining eight conserved master regulators of reprogramming-induced rejuvenation.
Significance. Partial reprogramming with OSKM can reverse molecular features of biological age but currently requires gene therapy delivery, with attendant safety, access, and regulatory constraints. If specific protein-protein interfaces, rather than enzymatic activities, drive the rejuvenation phenotype of reprogramming regulators, those interfaces become targetable by small molecules. A systematic panel of PPI biosensors for the nine conserved regulators would establish which interactions are causal, providing a target list for small-molecule rejuvenation drug discovery and replacing gene therapy as the modality required for therapeutic reprogramming. This project builds the foundational tool for one of the nine targets; demonstrating feasibility of the build-and-readout cycle is the prerequisite for scaling.
Yücel et al. (2025) reconstructed gene regulatory networks across partial reprogramming systems and identified conserved regulators associated with rejuvenation. A key observation motivating my project is that EZH2-Y726D, a catalytically impaired EZH2 mutant, can still support rejuvenation-associated effects. This suggests that at least some reprogramming regulators may have important non-canonical roles beyond their classic enzymatic activity.
PARP1 is another regulator in this general biological space. PARP1 is best known as a DNA damage response protein and poly(ADP-ribose) polymerase. Its catalytic activity uses NAD+ to build ADP-ribose chains on target proteins. However, PARP1 also participates in protein complexes, which makes it a good candidate for asking whether molecular interactions can be separated from enzymatic activity.
HPF1 stands for Histone PARylation Factor 1. It directly interacts with the PARP1 catalytic domain and changes how PARP1 modifies proteins. This makes HPF1 a useful partner for a simple biosensor: if PARP1 and HPF1 bind in the cell-free reaction, split GFP may reassemble and produce green fluorescence.
The PARP1-HPF1 interface has been structurally characterized by Suskiewicz et al. (2020) in Nature, who showed that HPF1 docks onto the PARP1 catalytic domain via a C-terminal peptide and completes the active site through an NAD+-independent composite interface. This makes PARP1-HPF1 an ideal first test case for a scaffolding-focused biosensor: the binding surface is structurally defined, the C-terminal region of HPF1 is the critical interaction element, which informs the N-terminal placement of GFP1-10, and the interaction is independent of PARP1 catalytic activity, allowing the WT versus E988K comparison to probe binding without confounding from enzymatic turnover.
GFP is the green fluorescent protein. In split-GFP systems, GFP is divided into two pieces:
If two proteins bring GFP1-10 and GFP11 close together, the GFP barrel can reassemble and become fluorescent. In my design, HPF1 carries GFP1-10 and PARP1 carries GFP11. Fluorescence therefore becomes a proxy for PARP1-HPF1 proximity.
The first aim of my final project is to build and test a cell-free split-GFP biosensor for the PARP1-HPF1 interaction by using DNA construct design, AlphaFold 3 structural validation of the fusion complex, Twist clonal gene synthesis, E. coli codon optimization, and Ginkgo Cloud Lab cell-free protein expression.
If the biosensor works, the next step is to add biochemical controls that distinguish binding from catalytic activity. This would require a PARP1 enzymatic activity assay, such as NAD+ depletion or PARylation detection, and expression quality control such as Echo-MS or SDS-PAGE.
The long-term vision is to create a panel of cell-free biosensors for conserved reprogramming regulators. Each biosensor would test a specific protein-protein or protein-DNA interaction and compare wild-type versus catalytic-dead or interaction-altered variants.
I ordered three clonal gene constructs from Twist Bioscience.
| Construct | Protein Design | Purpose |
|---|---|---|
| PARP1cat-WT-GFP11 | PARP1 catalytic domain, wild type, His6-tagged, C-terminal GFP11 | Positive PARP1 construct for HPF1 binding readout |
| PARP1cat-E988K-GFP11 | Same PARP1 catalytic domain, E988K mutation, His6-tagged, C-terminal GFP11 | First-pass mutant comparison |
| HPF1-GFP1-10 | Full-length HPF1, His6-tagged, N-terminal GFP1-10 | Binding partner and large split-GFP half |
Full-length PARP1 is large and multi-domain. Large human proteins can be difficult to express in E. coli cell-free lysate. I therefore use the PARP1 catalytic domain to make the construct more feasible for cell-free expression while keeping the region that interacts with HPF1.
HPF1 uses its C-terminal region to interact with PARP1. If I put the large GFP1-10 fragment on the C-terminus of HPF1, it might block the interaction I am trying to measure. Therefore, HPF1 is designed with GFP1-10 on the N-terminus.
E988 is part of the PARP1 catalytic machinery. The E988K mutant is expected to disrupt catalytic PARP activity. However, this project does not directly test catalytic activity. In this project, E988K is used as a first-pass mutant comparison in the biosensor.
The planned wet-lab assay uses Ginkgo Cloud Lab cell-free protein expression. Each reaction contains cell-free expression mix plus plasmid DNA. The key comparison is two-plasmid co-expression:
Controls are essential because GFP fluorescence can be misleading without them.
| Condition | Why It Matters |
|---|---|
| No DNA | Measures background fluorescence of the reaction |
| HPF1-GFP1-10 alone | Tests whether GFP1-10 gives signal by itself |
| PARP1cat-WT-GFP11 alone | Tests whether GFP11 gives signal by itself |
| PARP1cat-E988K-GFP11 alone | Same single-plasmid control for mutant |
| WT co-expression | Tests whether the biosensor works for the expected interaction |
| E988K co-expression | First-pass comparison against WT |
The direct readout is green fluorescence from reconstituted split GFP.
This score is useful as a first-pass comparison, but it must be interpreted carefully. A lower E988K signal could mean weaker binding, lower expression, worse folding, or worse split-GFP geometry.
This distinction is the most important part of the project. My claim is intentionally limited to the data this experiment can actually produce.
If the biosensor works, I expect the WT co-expression condition to produce fluorescence above the no-DNA and single-plasmid controls. That would mean the PARP1cat-GFP11 and HPF1-GFP1-10 fusion proteins can be expressed and can bring split-GFP fragments together.
For E988K, there are two useful outcomes:
Either result is useful, but neither result alone proves anything about cellular rejuvenation.
I validated the structural feasibility of the three-construct biosensor design by predicting the binary PARP1cat-GFP11 / HPF1-GFP1-10 complex using AlphaFold 3. This validates two design assumptions in a single prediction: (1) the fusion proteins fold without disrupting the PARP1-HPF1 binding interface, and (2) the split-GFP halves are brought into a geometry compatible with beta-barrel reassembly when the complex forms.
The validation used DNA construct design (assembly of fusion architectures in Benchling), protein design (placement of split-GFP halves and linkers to preserve the HPF1 C-terminal binding region), use of biological databases (UniProt, FPbase), and computational structure prediction (AlphaFold 3). Together these techniques convert a wet-lab design into a testable in silico model before any DNA is synthesized, allowing geometric and topological errors to be caught at design time rather than after Twist synthesis.
The AlphaFold 3 prediction returned ipTM = 0.71 and pTM = 0.64, indicating a confidently predicted inter-chain interface. The core regions of both chains, the PARP1 catalytic domain, the HPF1 globular core, and the reassembled split-GFP barrel, display pLDDT > 90 throughout. The PAE plot shows clean low-error blocks both along the diagonal (intra-chain confidence) and in the off-diagonal regions (inter-chain placement confidence). Disordered regions, comprising the N-terminal His-tags, the GSGSG and GSGSGSGSG linkers, and the C-terminal His-tag of HPF1, appropriately register as low pLDDT (orange). Qualitatively, the prediction shows the HPF1 C-terminal region docked at the PARP1 catalytic interface as expected from Suskiewicz et al. (2020), and the GFP1-10 and GFP11 fragments occupy adjacent positions consistent with beta-barrel reassembly. This is the strongest pre-wet-lab evidence achievable that the biosensor geometry supports the intended readout.
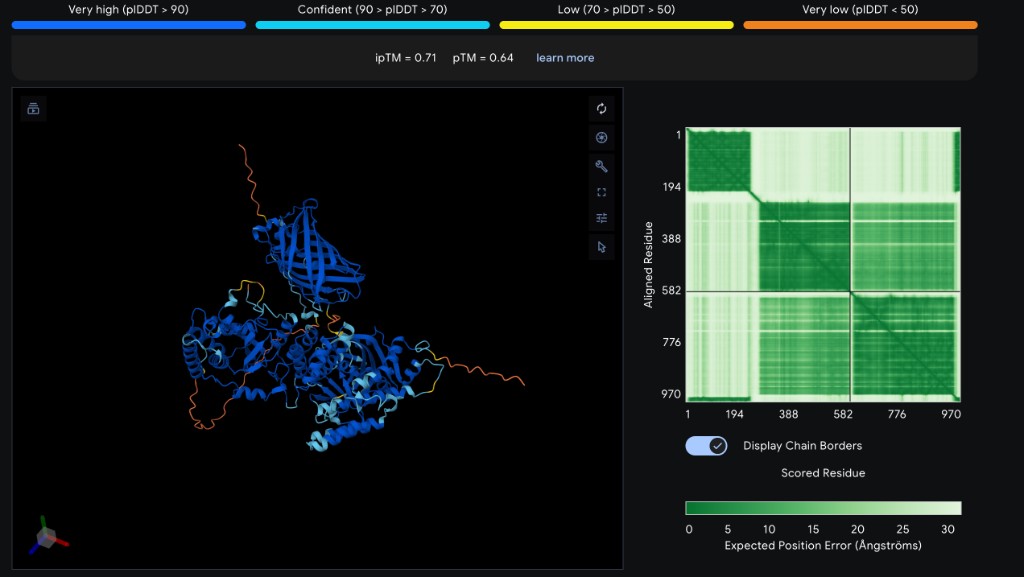
AlphaFold interpretation. Erstmal, das ist ein hervorragendes Ergebnis. ipTM = 0.71 ist solide (“confident interface”), pTM = 0.64 ist akzeptabel fuer ein 970-Residuen-Heterodimer mit flexiblen Tags, und das blaue Strukturzentrum zeigt klar zwei Dinge: (1) PARP1cat + HPF1 docken wie erwartet, (2) das Split-GFP-Barrel scheint im Komplex zu reassemblieren. Das ist genau die strukturelle Validierung des Biosensor-Designs.
Several challenges were resolved during the design-validation cycle and serve as informative findings. (1) Full-length BRCA1 was originally planned but exceeded the Twist Clonal Gene length limit of approximately 5 kb; this was resolved by restricting BRCA1 to the RING domain (aa 1-109) where C61G acts. (2) Initial vector selection was a CMV-promoter mammalian construct, which is incompatible with Ginkgo’s E. coli T7 cell-free system; this was resolved by switching to a pT7-blank Kan vector with a T7-RBS insertion site. (3) The original HPF1 fusion design placed GFP1-10 at the C-terminus, which would have occluded the HPF1 C-terminal PARP1-binding region; this was caught at the design-review stage and resolved by moving GFP1-10 to the N-terminus. (4) A residual risk remains: ipTM = 0.71 indicates good but not perfect interface confidence, leaving open the possibility that the fusion linkers introduce geometric strain not fully captured by the model, and that CFPS expression yields may differ between WT and E988K constructs in ways that confound the WT/E988K fluorescence comparison. This will be controlled empirically by single-plasmid normalization and Echo-MS expression QC on a subset of wells.
| Phase | Work | Expected Timing |
|---|---|---|
| Design | Finalize construct architecture and verify sequences | Completed |
| Build | Twist clonal gene synthesis and sequence verification | 1-2 weeks |
| Test | Ginkgo Cloud Lab cell-free expression and fluorescence readout | After constructs arrive |
| Analyze | Background correction, WT vs E988K comparison, figures | 1 week |
| Learn | Decide whether to improve tag placement, add controls, or scale to more regulators | Final project write-up |
This project has relatively low direct biosafety risk because it uses non-replicating cell-free reactions rather than engineered organisms released into the environment. The constructs encode human protein fragments and are intended for in vitro expression only.
The main ethical responsibility is truthful communication. Aging biology can easily be overhyped. I need to be clear that this project is not an anti-aging treatment, not a rejuvenation result, and not a clinical experiment. It is a molecular biosensor project that could support future mechanistic work.
Another ethical principle is non-maleficence: avoiding harm. In this context, harm could come from overstating weak evidence, especially in a field where people may be vulnerable to exaggerated longevity claims. I will therefore present the project as tool-building and clearly separate direct data from future speculation.
| Item | Approximate Cost | Notes |
|---|---|---|
| PARP1cat-WT-GFP11 clonal gene | Included in Twist order | ~1,170 bp |
| PARP1cat-E988K-GFP11 clonal gene | Included in Twist order | ~1,170 bp |
| HPF1-GFP1-10 clonal gene | Included in Twist order | ~1,743 bp |
| Twist total | ~$532.47 | 3 constructs |
| Ginkgo CFPS plate | TBD | Depends on HTGAA/Ginkgo pricing |
| Optional protein QC | TBD | Echo-MS or gel-based QC if available |
The project is intentionally small because the available budget is limited. A larger project testing all regulators would require many more constructs and assays.
If the biosensor works, the next steps are:
The strongest honest claim for this final project is:
I designed a three-construct, cell-free split-GFP biosensor for the PARP1-HPF1 interaction, validated the design in silico with AlphaFold 3 (ipTM = 0.71, recapitulated PARP1-HPF1 interface and split-GFP reassembly geometry), and ordered the constructs from Twist Bioscience for cell-free expression and plate-reader readout at Ginkgo Cloud Lab. This is a foundational HTGAA biosensor project: it produces an open, validated construct set that can be extended to the remaining eight conserved master regulators of reprogramming-induced rejuvenation, supporting the longer-term goal of identifying druggable PPI interfaces for small-molecule rejuvenation without gene therapy.