Week 11 HW: Bioproduction And Cloud Labs
HTGAA Week 11 Homework — Bioproduction & Cloud Labs
Part A: The 1,536 Pixel Artwork Canvas
What did you contribute to the community bioart project?
I contributed by correcting some of the pixel colours that appeared wrongly placed in the region slightly above and around the word “love” on the canvas, helping restore the intended colour arrangement in that section of the artwork.
What did you like about the project?
What I liked most was how a 1,536-well plate — a tool normally reserved for high-throughput biological experiments — became a shared creative medium connecting students across different institutions and time zones. It reframed what a cloud lab can do: not just automate experiments, but enable collective human expression at scale.
What could be made better for next year?
One improvement would be providing contributors with a live, real-time preview of the full canvas as it fills in, so each person can see the emerging collective image and make more intentional choices about colour and placement. A short window for reviewing and correcting misplaced pixels before the editing deadline closes would also improve the final artwork quality.
Part B: Cell-Free Protein Synthesis
Component Roles
Question: Referencing the cell-free protein synthesis reaction composition, provide a 1-2 sentence description of what each component’s role is in the cell-free reaction.
E. coli BL21(DE3) Star Lysate (includes T7 RNA Polymerase) Provides the core molecular machinery for transcription and translation, including ribosomes, tRNAs, aminoacyl-tRNA synthetases, elongation and release factors, and chaperones. The included T7 RNA Polymerase drives transcription specifically from T7 promoter-containing DNA templates.
Potassium Glutamate Supplies K+ ions required for ribosome assembly and optimal translation activity; glutamate is used as the counterion instead of chloride because it is less inhibitory to enzyme activity at physiological concentrations.
HEPES-KOH pH 7.5 A zwitterionic buffer that maintains the reaction pH near the physiological optimum (~7.5), ensuring consistent transcription and translation enzyme activity throughout the reaction.
Magnesium Glutamate Provides Mg2+, an essential cofactor for ribosome subunit assembly, mRNA binding, aminoacyl-tRNA accommodation, and nearly all NTP-dependent enzymatic steps in transcription and translation.
Potassium Phosphate Monobasic / Dibasic Functions as a secondary buffering system and provides inorganic phosphate (Pi), which participates in energy metabolism and nucleotide regeneration reactions.
Ribose A pentose sugar that feeds into nucleotide biosynthesis via the pentose phosphate pathway, supporting sustained NMP-to-NTP regeneration in long-duration reactions.
Glucose The primary carbon and energy source; drives ATP regeneration through glycolysis, sustaining the energy requirements of transcription and translation over time.
AMP, CMP, GMP, UMP Nucleoside monophosphates that serve as direct precursors for RNA synthesis; they are phosphorylated to their triphosphate forms by kinases retained in the lysate and incorporated during transcription.
Guanine A free nucleobase that enters GMP biosynthesis via the purine salvage pathway, supplementing the GMP pool without the cost of de novo synthesis.
17 Amino Acid Mix Provides the bulk of the 20 proteinogenic amino acids required for ribosomal translation; tyrosine and cysteine are supplied separately due to their individual handling requirements.
Tyrosine Supplied separately because of its very low solubility at neutral pH, which would cause precipitation if included in the master mix stock.
Cysteine Supplied separately because it is highly prone to oxidation; adding it fresh preserves its reduced thiol form, which is required for correct protein folding.
Nicotinamide A precursor to NAD+ that supports redox reactions in glycolysis and other metabolic pathways that regenerate energy for the reaction.
Nuclease-Free Water Used to bring the reaction to final volume; the absence of RNases and DNases protects the DNA template and mRNA transcript from degradation throughout the reaction.
Master Mix Comparison
Question: Describe the main differences between the 1-hour optimized PEP-NTP master mix and the 20-hour NMP-Ribose-Glucose master mix. (2-3 sentences)
The 1-hour PEP-NTP formulation uses phosphoenolpyruvate (PEP) as an energy regeneration substrate and pre-formed NTPs as direct transcription substrates, enabling rapid ATP regeneration via pyruvate kinase and immediate transcriptional activity — but PEP is consumed quickly and is relatively expensive, making this formulation best suited to short, high-intensity expression bursts. The 20-hour NMP-Ribose-Glucose formulation instead uses glucose (via glycolysis) and ribose (via the pentose phosphate pathway) to regenerate both energy and nucleotides from cheaper NMPs over time, creating a slower but far more sustained reaction. The tradeoff is a lower peak expression rate but higher cumulative yield, making it more appropriate for slowly-maturing fluorescent proteins or reactions requiring extended incubation.
Bonus Question
Question: How can transcription occur if GMP is not included but Guanine is?
The E. coli lysate retains active purine salvage pathway enzymes, most importantly hypoxanthine-guanine phosphoribosyltransferase (HGPRT), which catalyses the reaction: guanine + PRPP -> GMP + PPi, where PRPP is generated from ribose-5-phosphate. The resulting GMP is then phosphorylated to GDP and GTP by guanylate kinase and nucleoside diphosphate kinase, making GTP fully available for transcription. This approach is more cost-effective than supplying pre-made GMP while achieving the same functional outcome.
Part C: Planning the Global Experiment
Fluorescent Protein Properties
Question: Given the 6 fluorescent proteins used for the collaborative painting, identify and explain at least one biophysical or functional property of each protein that affects expression or readout in cell-free systems. (1-2 sentences each)
sfGFP sfGFP was engineered with mutations that dramatically improve folding robustness, making it resistant to aggregation even under the crowded, resource-limited conditions of a cell-free reaction — a key advantage over wild-type GFP. Like all GFP-family proteins, its chromophore maturation still requires molecular oxygen (O2) for the autocatalytic cyclization-oxidation reaction, so fluorescence output depends on adequate O2 availability during incubation.
mRFP1 mRFP1 has a notably slow and incomplete chromophore maturation rate (~1-2 hours to partial maturation at 37 degrees C) and relatively low quantum yield compared to newer red FPs. In a cell-free reaction, this means a significant fraction of translated mRFP1 protein may remain non-fluorescent at any given readout time point.
mKO2 mKO2 (monomeric Kusabira-Orange 2) has one of the slowest chromophore maturation times among common FPs (~4-5 hours for significant signal), and also shows moderate acid sensitivity, losing fluorescence at pH values below ~6.5. In a cell-free system, the long maturation time makes the 20-hour NMP-Ribose-Glucose formulation far more appropriate than the 1-hour PEP-NTP mix for detecting meaningful fluorescence signal.
mTurquoise2 mTurquoise2 has an exceptionally high quantum yield (~0.93, the highest reported for any cyan FP), making it highly bright and detectable even at low expression levels in cell-free reactions. However, like all GFP-family proteins it requires O2 for chromophore maturation, and it loses fluorescence under acidic conditions that may develop as the reaction consumes buffer capacity over time.
mScarlet-I mScarlet-I is notable for having one of the fastest maturation rates among red/orange fluorescent proteins (~45-60 minutes), which is a direct functional advantage in cell-free systems compared to slower red FPs like mRFP1 or mKO2. Its high brightness (high extinction coefficient combined with high quantum yield) makes it a sensitive readout even within the short 1-hour PEP-NTP reaction window.
Electra2 Electra2 is a biliverdin-dependent fluorescent protein whose chromophore is derived from the cofactor biliverdin rather than from autocatalytic amino acid cyclization. This gives it a critical advantage in cell-free reactions: chromophore maturation does not require molecular oxygen, unlike all GFP/RFP-family proteins — however, it requires biliverdin to be present in the reaction either from the lysate or supplied exogenously, making fluorescence output dependent on cofactor availability.
Reagent Optimization Hypothesis
Question: Create a hypothesis for how adjusting one or more reagents in the cell-free master mix could improve a specific biophysical or functional property you identified above, in order to maximize fluorescence over a 36-hour incubation. Clearly state the protein, the reagent(s), and the expected effect.
Protein: mKO2
Hypothesis: Because mKO2 suffers from slow chromophore maturation (~4-5 hours), a large fraction of translated protein remains non-fluorescent during the early phase of a 36-hour reaction. I hypothesise that supplementing the reaction with an additional 1-2 mM magnesium glutamate (above the standard concentration) will improve ribosome translational fidelity and elongation speed, increasing the total pool of correctly folded mKO2 protein available to enter the maturation pathway early in the reaction. With more correctly folded substrate accumulating simultaneously, total mature fluorescent mKO2 should reach a higher plateau faster, increasing both the rate of fluorescence accumulation and the final endpoint signal at 36 hours compared to the standard master mix formulation.
HTGAA Week 11 — Part C: Cell-Free Master Mix Design
Phase 2: Reagent Supplement Composition
Well Assignment
| Field | Detail |
|---|---|
| Quadrant | Q1 |
| Well Label | P14 |
| Fluorescent Protein | mKO2 |
| Node | USFQ Latin America (Quito, Ecuador) — Node 14 |
Hypothesis (from Phase 1)
mKO2 is one of the slowest-maturing fluorescent proteins in the panel, requiring approximately 4–5 hours to develop significant fluorescence. Because a large fraction of translated mKO2 protein remains non-fluorescent early in the reaction, maximising fluorescence over 36 hours requires increasing the rate and efficiency of correct protein folding before chromophore maturation can begin.
I hypothesised that supplementing the reaction with additional Magnesium Glutamate — above the standard master mix concentration — would improve ribosomal translational fidelity and elongation speed. This would increase the pool of correctly folded mKO2 protein available to enter the chromophore maturation pathway earlier, resulting in a higher fluorescence plateau at the 36-hour endpoint.
Reagent Supplement Applied
Based on my hypothesis, I made the following adjustment to the cell-free reaction for well Q1-P14:
| Reagent | Baseline Concentration | Adjusted Concentration | Change | Supplemental Volume |
|---|---|---|---|---|
| Magnesium Glutamate | 6.975 mM | 8.850 mM | +26.9% | 75 nL |
| Nuclease-Free Water | 2.000 µL | 1.925 µL | -3.7% | 1925 nL (auto-adjusted) |
All other reagents were left at their standard master mix concentrations. The total reaction volume remains 20 µL.
The Nuclease-Free Water was automatically reduced by the tool to compensate for the added Magnesium Glutamate volume, maintaining the correct total reaction volume. No other reagents were altered.
Reagent Supplement JSON
The following JSON was generated and saved by the rcdonovan.com/1536 platform upon submission:
Expected Outcome
By raising the Magnesium Glutamate concentration from 6.975 mM to 8.850 mM, I expect the ribosomes to function with greater fidelity and efficiency throughout the 36-hour incubation. This should result in:
- A larger pool of correctly folded mKO2 protein accumulating in the early hours of the reaction
- More protein entering the chromophore maturation pathway simultaneously
- A faster rate of fluorescence increase compared to wells using the standard master mix
- A higher total fluorescence endpoint at 36 hours
This prediction will be evaluated once fluorescence data is returned from the Ginkgo Nebula cloud lab.
Evidence Screenshots
Two screenshots were captured to document this submission:
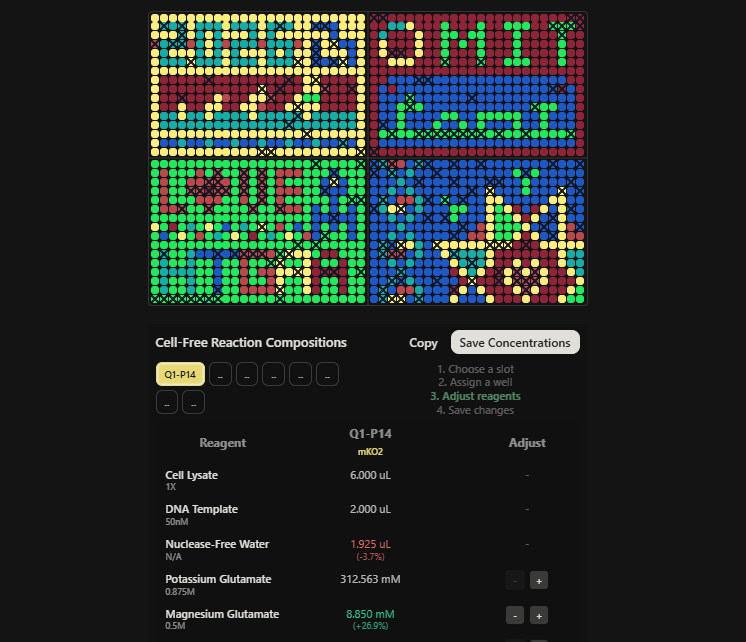
Screenshot 1 — HTGAA_W11_Q1P14_mKO2_reagent_composition.png
Shows the Cell-Free Reaction Compositions interface with well Q1-P14 assigned, mKO2 confirmed as the fluorescent protein, Magnesium Glutamate adjusted to 8.850 mM (+26.9%), and Nuclease-Free Water reduced to 1.925 µL (-3.7%).
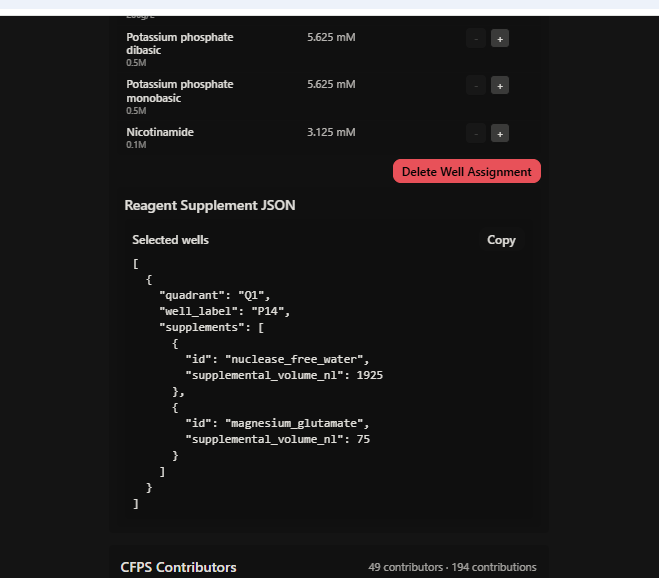
Screenshot 2 — HTGAA_W11_Q1P14_mKO2_JSON_saved.png
Shows the Reagent Supplement JSON successfully generated and the confirmation message: “Copied selected well supplement JSON to clipboard” — confirming the submission was saved to the platform.
Summary Statement
Well Q1-P14 (mKO2) was supplemented with 75 nL of additional Magnesium Glutamate, raising the final Mg²⁺ concentration by 26.9% to 8.850 mM. This single targeted adjustment was made within the 2 µL custom supplement volume and is directly motivated by mKO2’s known slow maturation kinetics.
References
Part A & B: Cloud Labs & Cell-Free Protein Synthesis Metabolism
Calhoun, K. A., & Swartz, J. R. (2005). Energizing cell-free protein synthesis with glucose and other carbohydrates. Biotechnology and Bioengineering, 90(5), 606–613. https://doi.org/10.1002/bit.20449
Caschera, F., & Noireaux, V. (2014). Synthesis of 2.3 mg/ml of green fluorescent protein in a cell-free system. Biochemical Engineering Journal, 94, 40–47. https://doi.org/10.1016/j.bej.2014.11.008
Jewett, M. C., Calhoun, K. A., Voloshin, A., Wuu, J. J., & Swartz, J. R. (2008). An integrated cell-free metabolic platform for protein synthesis and automated macromolecular assembly. Molecular Systems Biology, 4(1), 220. https://doi.org/10.1038/msb.2008.57
Sun, Z. Z., Hayes, C. A., Shin, J., Caschera, F., Murray, R. M., & Noireaux, V. (2013). Protocols for implementing an Escherichia coli based TX-TL cell-free expression system for synthetic biology. Journal of Visualized Experiments, (79), e50762. https://doi.org/10.3791/50762
Part C: Fluorescent Protein Properties & Biophysics
Bindels, D. S., Haarbosch, L., van Weeren, L., Postma, M., Wiese, K. E., Mastop, M., … & Gadella, T. W. (2017). mScarlet: A bright monomeric red fluorescent protein. Nature Methods, 14(1), 53–56. https://doi.org/10.1038/nmeth.4074
Goedhart, J., von Stetten, D., Noirclerc-Savoye, M., Lelimousin, M., Joosen, L., Hink, M. A., … & Gadella, T. W. (2012). Structure-guided evolution of cyan fluorescent proteins into a new class of highly efficient, monomeric FPs. Nature Communications, 3(1), 751. https://doi.org/10.1038/ncomms1738
Pédelacq, J. D., Cabantous, S., Tran, T., Terwilliger, T. C., & Waldo, G. S. (2006). Engineering and characterization of a superfolder green fluorescent protein. Nature Biotechnology, 24(1), 79–88. https://doi.org/10.1038/nbt1172
Rodriguez, E. A., Campbell, R. E., Lin, J. Y., Lin, M. Z., Miyawaki, A., Palmer, A. E., … & Tsien, R. Y. (2017). The growing and glowing toolbox of fluorescent and photoactive proteins. Trends in Biochemical Sciences, 42(2), 111–129. https://doi.org/10.1016/j.tibs.2016.09.010
Sakaue-Sawano, A., Kurokawa, H., Morimura, T., Re穩定, A., Hama, H., Shiba, H., … & Miyawaki, A. (2008). Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell, 132(3), 487–498. https://doi.org/10.1016/j.cell.2007.12.033