Week 7 Review: Genetic Circuits Part II
Week 7 — Genetic Circuits II: Neuromorphic Circuits & Fungal Materials
TL;DR: Cells can do more than switch genes on or off. By encoding signal weights in promoter and RBS strengths, and using RNA-cleaving enzymes as nonlinear activation functions, genetic circuits can implement perceptron-style neural computation — graded, multi-input, noise-averaging. This week also covers fungal materials: from mycelium composites and leather alternatives to engineering fungi as autonomous building repair agents. The worked DNA design demonstrates how to prepare a codon-optimised insert for two different assembly strategies.
Course: HTGAA Spring 2026 Lecture (Tue 17 Mar): Ron Weiss — Genetic Circuits Part II: Neuromorphic Circuits Recitation (Wed 18 Mar): Evan Holbrook, Ren Ramlan — Neuromorphic Circuits & Biomaterials Author: Fiona Connolly (Committed Listener BioPunk Node)
Why this matters
Week 6 showed how to build genetic circuits that compute Boolean logic — toggles, repressilators, AND gates. These are powerful, but they have a ceiling: they answer yes/no questions. Real biological decision-making is graded, not binary. A cell deciding whether to sporulate, a tumour deciding whether to metastasise, an immune cell deciding whether to activate — these are all weighted, multi-signal computations that binary gates cannot capture.
The Intracellular Artificial Neural Network (IANN) is an attempt to implement neural-network mathematics directly inside a cell. It is one of the most conceptually ambitious ideas in synthetic biology: repurposing the machinery of gene expression not as a logic gate but as an analog computer. At the same time, a completely different application of biological organisation — fungal mycelium — is showing that biology can build large-scale materials, not just molecular machines. Both threads converge on the same design philosophy: use the graded, adaptive properties of living systems rather than fighting them.
Core concepts
From Boolean to analog: why graded computation matters
A Boolean NOT gate needs a sharp threshold. If the input fluctuates around that threshold — as all biological signals do — the gate flips randomly. This is the threshold noise problem that limits digital genetic circuits.
An analog perceptron instead computes a weighted sum of multiple inputs:
output = f( w₁·x₁ + w₂·x₂ + … + wₙ·xₙ + b )
Because it integrates over several signals, random fluctuations in any single input are partially averaged out. The more inputs you sum, the better the noise averaging. This is why an IANN can be more robust than a Boolean gate even though its components are individually noisier — it exploits the statistics of biology rather than demanding perfect parts.
| IANNs | Boolean genetic circuits | |
|---|---|---|
| Output type | Continuous, graded | Binary (ON/OFF) |
| Multi-input handling | Weighted sum in one step — scales linearly | Gate cascades — grow exponentially with input number |
| Noise tolerance | Averaging across many inputs buffers fluctuations | Maximally sensitive to noise at the decision threshold |
| Classification power | Weighted patterns across many signals | Fixed logical conditions only |
| Biological relevance | Mirrors natural graded signalling | Imposes discrete logic on an analog biology |
| Implementation complexity | Hard — weights require careful promoter/RBS calibration | Mature toolkit, well-characterised parts |
| Single-cell reliability | Poor — population-level readout needed | Better for well-insulated designs |
The biological perceptron: translating mathematics into molecules
| Mathematical element | Biological implementation |
|---|---|
| Input xᵢ | Concentration of a transcription factor, RNA, or small molecule |
| Weight wᵢ | Promoter strength or RBS translation initiation rate |
| Weighted sum | Convergent transcriptional/translational co-regulation of one output mRNA |
| Bias b | Constitutive basal expression from a leaky promoter |
| Activation function f | A nonlinear molecular switch — typically a threshold-dependent RNA-cleaving enzyme |
Csy4: the molecular activation function
Csy4 (Cas6f) is a CRISPR-associated RNA endoribonuclease from Pseudomonas aeruginosa. It cleaves RNA at a specific 28-nucleotide hairpin with high fidelity and is orthogonal to most cellular RNA-processing machinery in E. coli and mammalian cells.
In an IANN, Csy4 serves two roles simultaneously: it is the output of one computational layer (its concentration is the layer’s result) and the activation function of the next (its threshold concentration creates the nonlinearity needed for meaningful computation).
In the canonical forward-activation topology used by the Weiss lab and depicted in the circuit diagram below: a structured 5′ hairpin engineered into the target mRNA occludes the ribosome binding site, suppressing translation in the absence of Csy4. When Csy4 accumulates above threshold, it cleaves the hairpin and releases the RBS, enabling translation. Csy4 is therefore an activator in this design. (An alternative repression topology — where cleavage removes an mRNA-stabilising element and accelerates decay — exists in principle but is less commonly deployed and is not the topology shown here.)
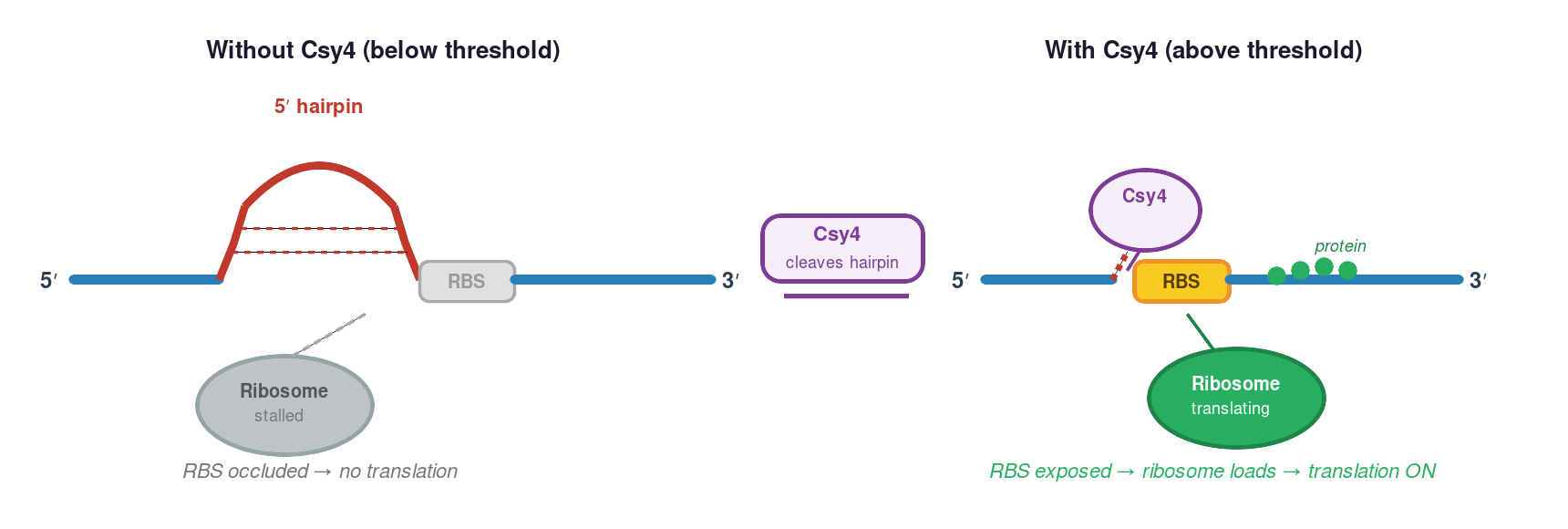
Key requirement: If Csy4 is Layer 1’s output, Layer 2 must use a different orthogonal endoribonuclease (a distinct Cas6 family member with a different recognition hairpin). Without orthogonality, Layer 1’s enzyme inadvertently regulates Layer 2’s targets and the layer separation collapses.
Is a reliable biological perceptron achievable?
Honestly: at the single-cell level, no — not yet. Gene expression is inherently stochastic (transcriptional bursting, ribosome noise, extrinsic variation in polymerase and ribosome concentrations). Single-cell IANN outputs are noisy.
At the population level, the answer is more encouraging. Analog computation averages over many noisy cells, and published IANNs demonstrate meaningful classification accuracy at the population level. The tradeoff is that single-cell sensing applications remain out of reach for now.
Strategies to reduce noise in IANN designs:
| Strategy | Effect |
|---|---|
| Chromosomal integration (vs. plasmid) | Eliminates copy-number variation — the largest source of extrinsic noise |
| Strong constitutive promoters for weight-encoding genes | Pushes into high-copy regime where coefficient of variation is lower |
| Insulated genetic parts (strong terminators flanking each cassette) | Prevents read-through from altering expression levels |
| More inputs per layer | Central-limit averaging — noise decreases as 1/√N inputs |
| Population-level readout | Exploit the statistics directly |
Parts libraries for IANN design
No unified IANN-specific parts library exists yet — the field is roughly 5–7 years behind digital genetic circuits in parts infrastructure. The practical toolkit today:
- Anderson Promoter Library (iGEM Registry: BBa_J23100–J23119) — 20 constitutive E. coli promoters spanning ~3 orders of magnitude in strength. The go-to resource for weight encoding.
- Salis Lab RBS Calculator (rbs.psu.edu) — predictive tool for designing RBS sequences to a target translation rate. Accuracy ~2–3 fold, sufficient for coarse weight setting.
- Csy4 and orthogonal Cas6 variants — recognition hairpin characterised by Haurwitz et al. (2010); adapted as a post-transcriptional multiplexer in mammalian gene circuits by Nissim et al. (Molecular Cell, 2014; PMID 24837679). Multiple orthogonal Cas6-family members now available for multilayer designs.
- Cello (Nielsen et al., 2016) — the gold standard for digital genetic circuit design automation; not IANN-specific, but the benchmark to understand what the digital alternative offers.
Multilayer IANN: circuit topology
The diagram below shows a two-layer IANN. X₁ drives Csy4 expression (Layer 1 component). Csy4 regulates Endo2 translation via a 5′ hairpin (Layer 1 output). Endo2 in turn regulates fluorescent protein translation via a second, orthogonal 5′ hairpin (Layer 2 output).
flowchart LR
subgraph L1["LAYER 1"]
X1["X₁ DNA\n(Csy4)"] -->|Tx| cm["Csy4 mRNA\n(no hairpin)"]
cm -->|Tl| CSY4["Csy4\nEndoribonuclease"]
X2["X₂ DNA\n(Endo2)"] -->|Tx| em["Endo2 mRNA\n⌇ 5′ hairpin ⌇"]
CSY4 -.->|"cleaves hairpin\n(regulates Tl)"| em
em -->|Tl| ENDO2["Endoribonuclease 2\n◄ Layer 1 OUTPUT"]
end
subgraph L2["LAYER 2"]
X3["X₃ DNA\n(FP)"] -->|Tx| fm["FP mRNA\n⌇ 5′ hairpin ⌇"]
ENDO2 -.->|"cleaves hairpin\n(regulates Tl)"| fm
fm -->|Tl| FP["Fluorescent Protein\n◄ Layer 2 OUTPUT"]
end
ENDO2 -->|"bridges L1 → L2"| L2Tx = transcription; Tl = translation; dashed arrows = post-transcriptional regulation via hairpin cleavage. Solid borders = constitutively expressed. Dashed borders = regulated mRNAs.
Reading the circuit: The concentration of Csy4 protein (Layer 1) is a continuous, weighted function of X₁ input strength. Above threshold, Csy4 enables (or represses) Endo2 translation. Endo2 concentration is the Layer 1 output — it is passed forward and determines fluorescent protein expression in Layer 2. The final FP signal is a nonlinear, two-layer function of all three DNA inputs.
Worked example: prostate cancer multi-biomarker IANN
A two-layer IANN is well matched to the prostate cancer monitoring problem. No single marker (PSA, PCA3, AR-V7) is reliably specific — but their weighted combination is far more informative. The IANN also needs to distinguish a high-priority metastasis alert from a medium-priority inflammatory warning, which requires two parallel output arms with different thresholds.
Circuit architecture
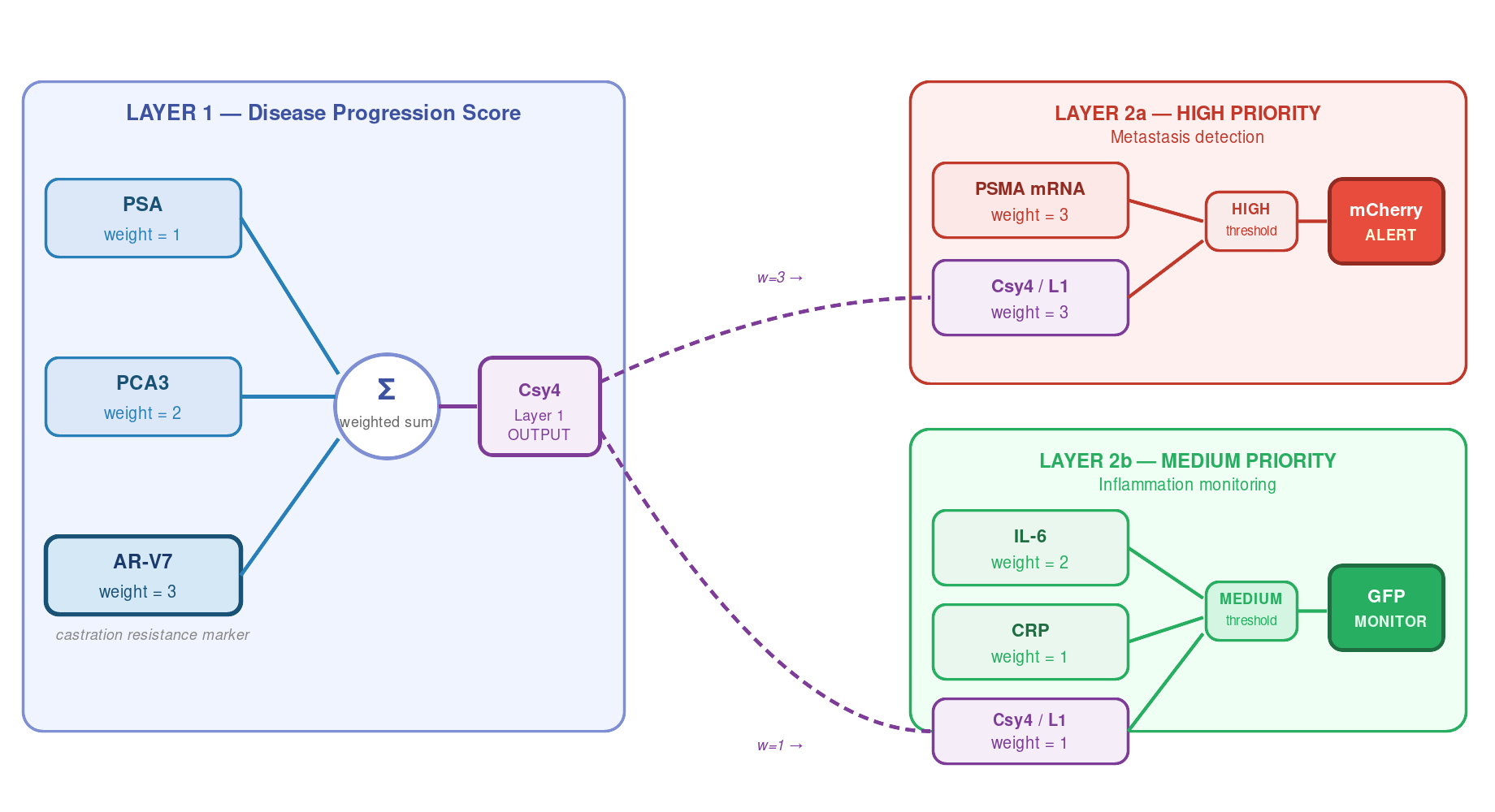
Biomarker rationale
| Layer | Biomarker | Significance | Weight |
|---|---|---|---|
| 1 | PSA | Prostate cancer progression; poor specificity alone | 1 |
| 1 | PCA3 | Prostate-specific non-coding RNA; more specific than PSA | 2 |
| 1 | AR-V7 | Androgen receptor splice variant — castration-resistant prostate cancer; predicts resistance to enzalutamide/abiraterone | 3 |
| 2a | PSMA | Prostate-specific membrane antigen; highly overexpressed in metastatic disease; FDA-approved PET imaging target | 3 |
| 2a | Csy4 (from L1) | Recurrence/CRPC score passed forward | 3 |
| 2b | IL-6 | Pro-inflammatory cytokine; drives STAT3/JAK signalling; promotes castration resistance at high levels | 2 |
| 2b | CRP | Systemic inflammation marker; elevated CRP is independently prognostic in metastatic disease | 1 |
| 2b | Csy4 (from L1) | Disease-driven vs. infection-driven inflammation disambiguation | 1 |
Tiered output interpretation
| mCherry (L2a) | GFP (L2b) | Clinical interpretation | Priority |
|---|---|---|---|
| OFF | OFF | No significant signal | Baseline monitoring |
| OFF | ON | Inflammation elevated, disease markers low — likely infection | Medium — investigate |
| ON | OFF | Metastatic markers elevated without systemic inflammation — early spread | High — immediate workup |
| ON | ON | Metastasis confirmed + dangerous inflammation | Critical |
Limitations
Sensing mechanism gaps. AR-V7 is a splice isoform of androgen receptor mRNA. Detecting it inside a living cell requires distinguishing the unique exon 3 / cryptic exon 3b junction sequence from full-length AR mRNA — not simply sensing AR expression. Toehold switches targeting the junction sequence could theoretically accomplish this, but no characterised part exists in any registry. PCA3 is a long non-coding RNA (lncRNA) that is predominantly nuclear-retained; most cytoplasmic RNA-sensing architectures (riboswitches, toehold switches) cannot access it — this is a compartmentalisation barrier, not merely a parts scarcity problem. PSMA (Layer 2a, w=3) is a cell-surface transmembrane protein, not a diffusible cytoplasmic molecule. An intracellular circuit cannot sense it directly; sensing would require reading PSMA mRNA levels via a toehold switch, or redesigning Layer 2a around a soluble marker with equivalent clinical specificity for metastatic disease.
Fixed weights. Weights are fixed at fabrication time — personalising thresholds for individual patients (e.g. those with baseline BPH-elevated PSA) requires re-engineering the promoter/RBS combination, which is not scalable in current form. The weight assignments shown (PSA w=1, PCA3 w=2, AR-V7 w=3, etc.) are illustrative choices made for didactic clarity, not values calibrated to clinical sensitivity/specificity data.
Population readout. Single-cell stochasticity means this would function as a population-averaged ex vivo diagnostic assay, not a single-cell in vivo sensor.
Fungal materials: biology at the macro scale
Fungi produce useful materials across three categories: structural composites, textiles, and food proteins.
Existing fungal materials
Mycelium composites — Ecovative / grow.bio, Ganoderma and Pleurotus ostreatus. Agricultural waste (corn husks, hemp hurds) inoculated with mycelium; the fungal network binds substrate into rigid, foam-like blocks for packaging and insulation. Competing with expanded polystyrene: compostable and carbon-negative, but moisture-sensitive (chitin is hygroscopic) and slower to produce at scale.
Fungal leather — MycoWorks (Reishi™ / Fine Mycelium™) and Bolt Threads (Mylo™), both Ganoderma-based. Dense mycelial mats processed into textile resembling full-grain leather. Attracted major fashion collaborations (Stella McCartney, Hermès, Adidas). Important 2025–2026 update: MycoWorks entered insolvency in October 2025 and was liquidated; Bolt Threads also ceased operations. Their failure is itself a data point — fungal materials face a difficult valley between proof-of-concept and commercial scale, and the funding environment for novel biomaterials contracted sharply in 2024–2025.
Mycoprotein — Quorn, Fusarium venenatum. Continuous fermentation on glucose, harvested and heat-treated (to reduce RNA content and gout risk), textured into meat analogues. ~45% protein by dry weight, complete amino acid profile, substantially lower land, water, and GHG footprint than animal protein. The most commercially mature fungal material category, launched in 1985, now in mainstream retail.
| Material | Species | Link | Traditional counterpart | Key advantage | Key disadvantage |
|---|---|---|---|---|---|
| Mycelium composite | Ganoderma, Pleurotus | Ecovative | Expanded polystyrene | Compostable, waste feedstock | Moisture-sensitive; cost gap |
| Fungal leather | Ganoderma | MycoWorks (liquidated 2025) | Animal leather | No slaughter; tunable texture | Needs polymer finish; companies failed at scale |
| Mycoprotein | Fusarium venenatum | Quorn | Meat / animal protein | Lower emissions; complete protein | Allergic reactions in some consumers |
Why fungi for synthetic biology?
The core argument: choose fungi when your protein requires eukaryotic processing, when secretion titre matters, or when the biomass itself is the product. Choose bacteria when speed, genetic precision, or parts-library depth are the priority.
| Criterion | Fungi | Bacteria (E. coli, B. subtilis) |
|---|---|---|
| Post-translational modifications | Full eukaryotic glycosylation, disulfide bonds | None — proteins misfold or form inclusion bodies |
| Secretion capacity | Exceptional — A. niger secretes >100 g/L enzyme industrially | Poor — most protein stays intracellular |
| Substrate flexibility | Grow on lignocellulosic waste | Require refined carbon sources |
| Biomass as product | Mycelial network IS the material | Biomass not structurally useful |
| Existing infrastructure | Centuries of industrial fermentation | Shorter history at large scale |
| Genetic toolkit | Improving (CRISPR in Aspergillus, Yarrowia) but behind bacteria | Largest and most mature in all of biology |
| Growth rate | Hours to days per doubling | ~20 min (E. coli) |
| Multi-nuclearity | Multiple nuclei per hypha — harder clean edits | Single chromosome, haploid — clean knockouts straightforward |
Worked example: engineering fungi for self-healing buildings
The long-term vision is a building material that behaves like living tissue — binding itself together during construction, detecting damage, and repairing autonomously without human intervention. Genetically engineered fungal mycelium, pre-incorporated as dormant spores, could fulfil all three roles.
flowchart LR
subgraph P1["① BIND"]
A["Spores mixed\ninto aggregate"] --> B["Mycelium threads\nthrough gaps"]
B --> C["Living composite:\ntensile + compressive\nstrength"]
end
subgraph P2["② DETECT"]
D["Crack opens"] --> E["Local O₂↑ CO₂↑\nmoisture↑"]
E --> F["Chemotropic growth\ntoward damage zone"]
end
subgraph P3["③ REPAIR"]
G["Urease overexpressed\n→ MICP: CaCO₃ fill"] --> H["Silk-like proteins\nsecreted → tensile matrix"]
H --> I["Composite repair:\nmineral + fibre"]
end
P1 -->|"Material ages\nor stressed"| P2
P2 -->|"Mycelium reaches crack"| P3MICP (Microbially Induced Calcite Precipitation): overexpressed urease converts urea → NH₃ + CO₂, raising local pH and precipitating CaCO₃ crystals that fill the crack void. The mycelial network simultaneously provides tensile reinforcement across the crack faces — a composite repair rather than a brittle mineral plug.
The Fungiteria model (a proposed fungi-bacteria consortium concept, coined in the self-healing materials research context; not yet standardised terminology) extends this further: bacteria (Sporosarcina pasteurii, the workhorse of bacterial MICP) live inside the mycelial network, performing high-efficiency mineralisation while the fungus provides macroscale navigation and scaffolding — a division of labour neither organism achieves alone.
Engineering targets
| Target | Implementation | Purpose |
|---|---|---|
| Crack chemotropism | Overexpress O₂/CO₂ gradient receptors | Directs growth to damage zone |
| MICP | Overexpress urease + carbonic anhydrase | CaCO₃ mineral fill |
| Alkaline tolerance | Engineer pH 12-tolerant spore germination | Concrete is pH 12–13 |
| Structural protein secretion | Secrete silk-like or elastin-like proteins | Tensile reinforcement of repair matrix |
| Dormancy control | Robust sporulation; germination triggered by crack signals | Lifetime persistence in material |
Why fungi over bacteria alone?
Bacterial self-healing concrete (Jonkers HM et al., Ecological Engineering 2010; 36:230–235; doi:10.1016/j.ecoleng.2008.12.036) already exists and works for microscale cracks. Fungi add what bacteria cannot provide:
| Criterion | Fungi | Bacteria (Bacillus MICP) |
|---|---|---|
| Growth scale | Centimetres — relevant to structural cracks | Confined to microscale voids |
| Mechanical role | Mycelial fibre network provides tensile reinforcement | Mineral fill only |
| Binding during fabrication | Can bind aggregate as living composite | Cannot serve as structural binder |
| Complex protein secretion | Correctly folded silk/elastin structural proteins | Cannot produce glycosylated or disulfide-bonded proteins |
Limitations
Concrete alkalinity (pH 12–13) is the primary biological barrier — most fungi cannot survive it, and engineering alkali-tolerant germination from scratch remains unsolved. Nutrient supply for long-term mycelial activity must be pre-loaded into the material, creating a finite repair budget. An engineered strain growing in a building must not sporulate and disperse to the environment — kill switches or strict auxotrophy are required. No regulatory pathway currently exists for living engineered organisms as structural building components.
Part 3: First DNA Twist order — CotB insert design
Assignment context: Design at least one insert sequence for the first Twist order. Per HTGAA TA guidance for committed listeners, this is a placeholder submission to confirm the final project direction. The sequence chosen is relvevant to my final project protein — B. subtilis CotB spore coat protein — codon-optimised for B. subtilis, nanobody-fusion-ready (no stop codon).
Protein and sequence overview
CotB is a B. subtilis spore coat protein used for surface display. The nanobody will be fused at the C-terminus of CotB, displayed on the spore surface. No stop codon is included in the insert so that the reading frame continues into the nanobody.
| Property | Value |
|---|---|
| Protein | CotB — B. subtilis spore coat protein |
| Length | 372 amino acids |
| DNA insert | 1116 bp, codon-optimised for B. subtilis |
| Stop codon | None (nanobody fusion at C-terminus) |
| GC content | 36.9% (within Twist synthesis range 25–65%) |
| Internal BsaI sites | 0 (clean for BsaI Golden Gate without domestication) |
| Backbone vector | pDG1730 (amyE integration, spectinomycin resistance) — working vector |
Two assembly versions
Version 1 — Gibson Assembly (1116 bp bare insert)
The CotB ORF is ordered from Twist as-is. Overlaps to the backbone (20–25 bp) are added at cloning time via PCR primers — the insert sequence itself requires no modification. The reverse primer tail should overlap into the start of the nanobody/linker sequence in the linearised backbone.
Version 2 — BsaI Golden Gate (1138 bp full synthesised fragment)

After BsaI digestion:
- Left 5′ overhang:
ATGA— CotB nt 1–4; standard ATG start - Right 5′ overhang:
GAAA— CotB nt 1113–1116; Lys C-terminus; junction with nanobody/linker insert
After ligation, the full 1116 bp CotB ORF is reconstructed intact with no insertion or deletion at either junction. The right overhang GAAA must match the left overhang on the downstream nanobody/linker piece — confirm this when designing that insert.
⚠️ Note on the right overhang:
GAAAis AT-rich (25% GC). If ligation efficiency is suboptimal, consider extending CotB by 1–2 codons to achieve a more GC-balanced overhang junction.
Both sequences are in COTB_Sequences.fasta (updated 2026-05-23), with full QC documentation in notes.md.
Pitfalls, controls, and how to know it worked
For IANN designs:
- Threshold miscalibration. If the activation-function threshold (Csy4 concentration for cleavage) is set too high or too low, the perceptron will always output ON or always OFF regardless of inputs. Titrate Csy4 expression level with a panel of RBS strengths and validate the transfer curve (input concentration vs. output) before full circuit assembly.
- Cross-talk between layers. If Csy4 accidentally cleaves the Layer 2 hairpin (orthogonality failure), Layer 1 computation bleeds directly into Layer 2 output. Test each endoribonuclease in isolation against every hairpin in the circuit before assembly.
- Plasmid copy-number noise. Large cell-to-cell variation in plasmid copy number creates extrinsic noise that dominates IANN output variation. Chromosomally integrate the circuit for reproducible results; validate by comparing plasmid vs. integrated output distributions.
- Control: single-input circuits (one xᵢ at a time) should produce a monotonic transfer curve. Non-monotonic behaviour indicates cross-regulation or unexpected feedback.
For fungal materials and engineering:
- Contamination. Fungal cultures are highly susceptible to faster-growing mould contamination. Work in controlled humidity and temperature; test on sterilised substrate before scaling.
- Multi-nuclearity. Transformed nuclei may be diluted by wild-type nuclei in the same hypha (heterokaryon). Maintain selection pressure throughout propagation; verify homokaryon status by single-spore isolation before characterisation.
- Moisture sensitivity. Mycelium composites lose structural integrity when wet. Measure compressive strength before and after water immersion — if values diverge >30%, the material is not suitable for structural applications without additional treatment.
For the CotB insert/Twist order:
- Confirm insert sequence by Sanger sequencing after cloning; verify the reading frame continues correctly into the nanobody fusion.
- Verify surface display by immunofluorescence with an anti-nanobody antibody on spores before functional assays.
Recommended reading
Daniel R, Rubens JR, Sarpeshkar R, Lu TK. Synthetic analog computation in living cells. Nature 2013; 497:619–623. doi:10.1038/nature12148 The foundational demonstration that analog gene circuits can compute logarithmic, additive, and ratiometric functions in living E. coli using just three transcription factors.
Pandi A, Koch M, Voyvodic PL et al. Metabolic perceptrons for neural computing in biological systems. Nature Communications 2019; 10:3880. doi:10.1038/s41467-019-11889-0 Implements four-input perceptrons for binary classification of metabolite combinations in whole-cell and cell-free systems — the closest published equivalent to a biological neural network classifier.
Nielsen AAK, Der BS, Shin J et al. Genetic circuit design automation. Science 2016; 352:aac7341. doi:10.1126/science.aac7341 Cello: automated design of Boolean genetic circuits from Verilog code. Essential baseline for understanding what the digital-circuit alternative offers — and why analog IANNs are architecturally distinct.
Cerimi K, Akkaya KC, Pohl C et al. Fungi as source for new bio-based materials: a patent review. Fungal Biology and Biotechnology 2019; 6:17. doi:10.1186/s40694-019-0080-y Systematic survey of mycelium-based materials patents 2009–2018 across packaging, textiles, insulation, and fire protection. Maps the commercial landscape and identifies the open engineering challenges.
Course resources
HTGAA 2026 course page — Week 7:
https://2026a.htgaa.org/2026a/course-pages/weeks/week-07/index.htmlLecture: Ron Weiss (MIT) — Genetic Circuits Part II: Neuromorphic Circuits
Ecovative / grow.bio: https://grow.bio — mycelium materials, GIY kits
Quorn mycoprotein science: https://www.quornnutrition.com/mycoprotein/what-is-mycoprotein
Bhattacharya et al. (2024) — Biomolecular ANN review (HTGAA assigned reading): doi:10.1016/j.biosystems.2024.105164
Anderson Promoter Library (iGEM): http://parts.igem.org/Promoters/Catalog/Anderson
Salis Lab RBS Calculator: https://salislab.net/software/
CotB sequences (this project):
COTB_Sequences.fasta— Gibson + BsaI Golden Gate inserts