Week 11 Review: Bioproduction & Cloud Labs
Week 11 — Bioproduction & Cloud Labs
One-line takeaway. A cloud lab is a wet-lab you drive from a laptop. This week you design a cell-free protein synthesis (CFPS) reaction that will run on one, in a global 1,536-well bioart canvas.
| Course | HTGAA Spring 2026 |
| Lecture | Tues, Apr 14, 2026 — Reshma Shetty, Bioproduction & Cloud Labs |
| Recitation | Wed, Apr 15 — Ronan Donovan, Cloud laboratories |
| Author | Fiona Commited Listener BioPunk SF |
Why CFPS as the canonical case for cloud labs?
A cell-free reaction is the fastest way to make protein. Make protein, read out a signal, learn something, design the next round — that loop is what synthetic biology runs on. The faster the loop, the more design space you get to search.
Cloud labs let you run that loop without owning a lab. You write a protocol, queue it, and a fleet of robots executes overnight. That changes who can do science (you, on a Tuesday) and how reproducible it is (every parameter is explicit because a robot needs it explicit).
Tie-ins to your final projects:
- MS2 L-protein (group project) — dozens of designed mutants, identical expression-and-readout pipeline. A textbook cloud-lab use case.
- Cholera Shield (individual final project) — Week 9 anti-cholera-toxin module will need cell-free expression conditions; the master-mix logic locked this week is the foundation.
Vocabulary (skim once, refer back later)
| Term | What it is |
|---|---|
| Cloud lab | A wet-lab facility you drive remotely. Ginkgo Nebula, Emerald, Strateos. |
| RAC | Reconfigurable Automation Cart — one instrument + a robotic arm + a software wrapper. |
| CFPS | Cell-free protein synthesis. Protein made in a tube of E. coli extract; no living cells. |
| Lysate | The soluble fraction of broken-open E. coli — ribosomes, tRNAs, enzymes. Source of all the machinery. |
| BL21 (DE3) Star | The standard CFPS strain. (DE3) = carries T7 RNA polymerase. Star = truncated RNase E (rne131), so mRNA lasts longer. |
| T7 promoter | TAATACGACTCACTATAGGG — the canonical 20-bp T7 RNAP cassette (–17 to +3), including the GGG initiator triplet required for efficient transcription start. Why your plasmid has one. |
| NTP / NMP / NDP | Nucleotide tri- / mono- / di-phosphates. NTPs are the working currency; NMPs are the cheap precursor. |
| Master mix | Pre-mixed cocktail of everything in the reaction except lysate and DNA. Used at 2× and diluted into the well. |
| Maturation half-time | How long after translation finishes before the chromophore lights up. Dominant property for CFPS readout. |
How a cloud lab is built
Reshma’s mental model is a 2×2 of automation vs flexibility:
quadrantChart
title Lab work, on two axes
x-axis "Low flexibility" --> "High flexibility"
y-axis "Low automation" --> "High automation"
quadrant-1 "Autonomous lab (Waymo)"
quadrant-2 "Work cell (red line)"
quadrant-3 "—"
quadrant-4 "Lab bench (car)"
"Lab bench": [0.85, 0.15]
"Work cell": [0.15, 0.85]
"Walk-up Opentrons": [0.5, 0.5]
"Cloud lab (Nebula)": [0.85, 0.85]Lab bench = max flexibility, near-zero automation. Reshma’s framing: the vast majority of research budgets — both academic and pharma R&D — still sit here. Work cell = max automation, locked to one workflow. Cloud lab = both. The hard engineering problem.
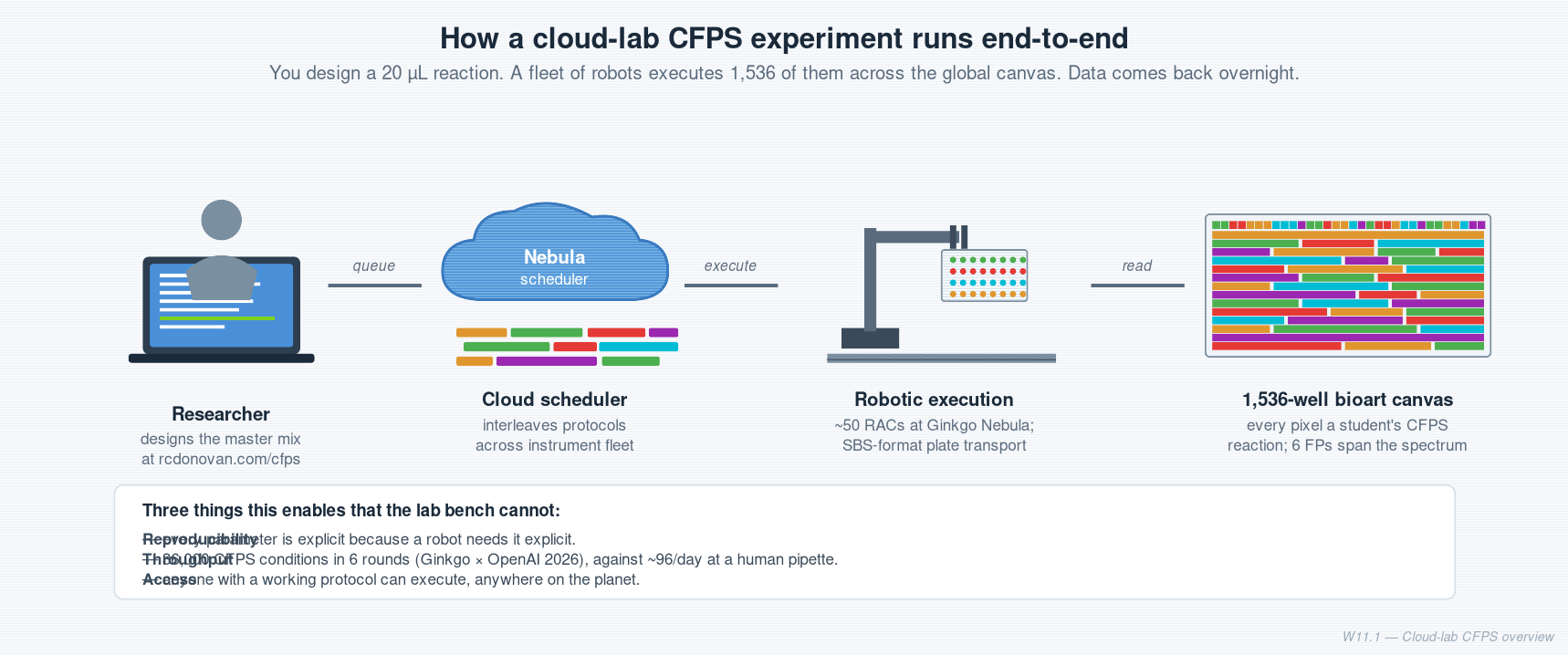
Fallback rendering. If the quadrant chart above doesn’t render in your viewer, here’s the same information as a table:
Mode Flexibility Automation Example Lab bench High Low A graduate student pipetting Walk-up automation Medium Medium Opentrons in the HTGAA teaching lab Work cell Low High NGS sample-prep robot Cloud lab High High Ginkgo Nebula, Emerald Cloud Lab
What’s inside a Nebula rack
Each rack is one instrument inside a standardized enclosure, with a robotic arm and a sample transport track running past it.
flowchart LR
subgraph RAC["A single RAC"]
Inst[Instrument<br/>e.g. centrifuge, Echo, Bravo, reader]
Arm[Robotic arm]
SW[Software wrapper<br/>parameterized control]
end
Track[Sample transport track<br/>SBS-format plates]
Plate[Plate]
Plate --> Track
Track <--> Arm
Arm <--> Inst
Inst <--> SWA few practical points worth knowing:
- ~50 racks in Boston right now, targeting 100.
- ~2 months to onboard a new instrument (custom internal fittings — “iKit” — for each one).
- SLAS / SBS plate format is the only meaningful adopted standard in the industry.
- Centrifuges become bottlenecks before mass specs. Because every protocol “quick-spins” everything.
- Nebula exposes
Generic_*modules (Generic_echo_hitpick,generic_multiflo_dispense,generic_spark_read, etc.) rather than packaged workflows. You compose your own.
Design choice worth noting. Ginkgo is building purpose-built automation hardware, not humanoid robots. Reshma is open that this is a bet. Other labs are taking the opposite bet.
Cell-free protein synthesis — what’s in the tube
A CFPS reaction is a protein factory in a tube. The factory needs continuous energy, building blocks, the right ionic environment, and the machinery to run it. The reagents map cleanly onto those four roles:
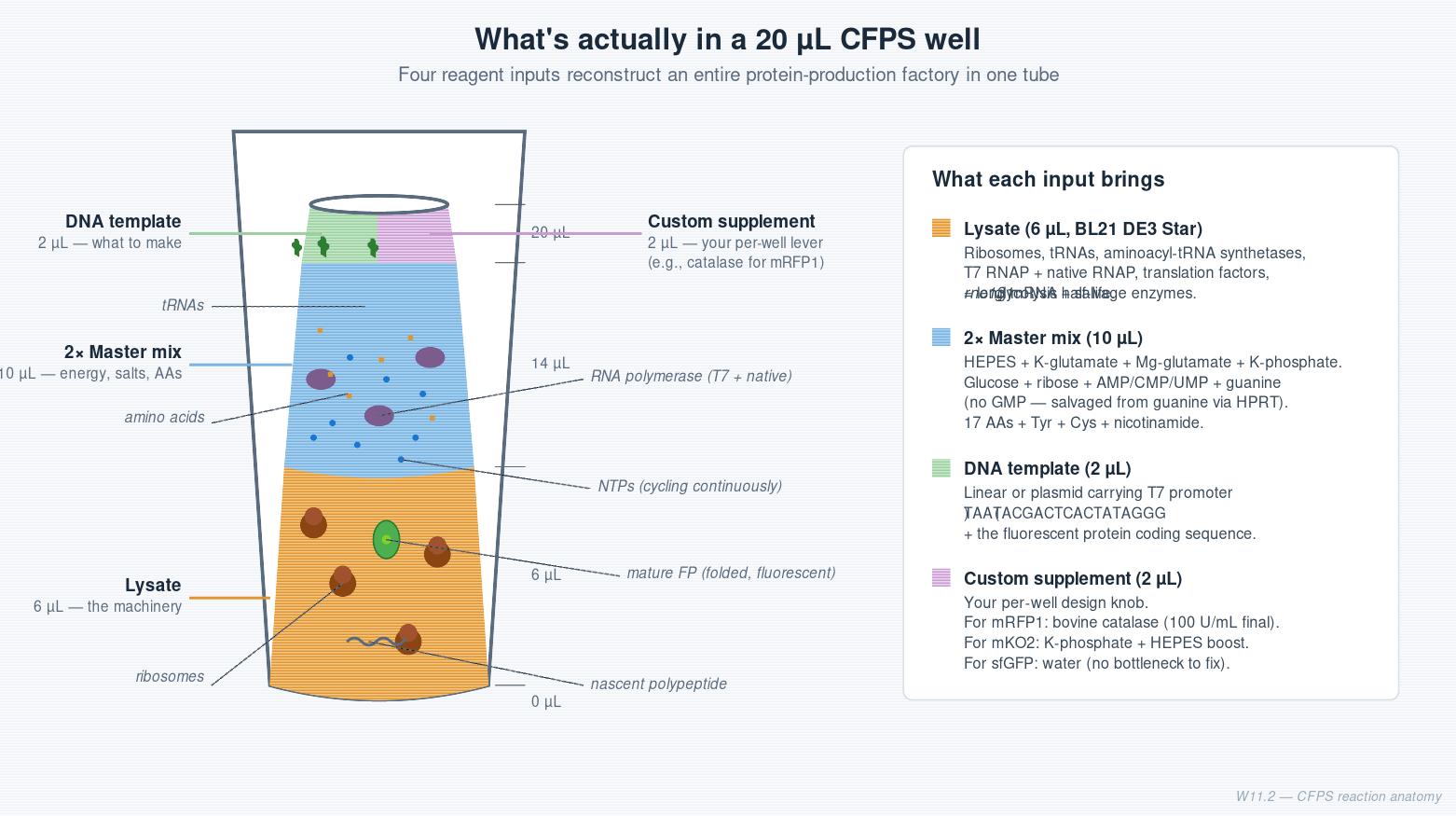
| Role | Reagents | What they do |
|---|---|---|
| Machinery | BL21 (DE3) Star lysate | Ribosomes, tRNAs, RNA polymerase, all translation factors, metabolic enzymes. T7 RNAP is pre-induced before lysis. Star = rne131 truncated RNase E → longer mRNA half-life. |
| Ionic environment | K-glutamate (130 mM), HEPES-KOH pH 7.5 (50 mM), Mg-glutamate (10 mM Mg²⁺), K-phosphate mono + dibasic (10 mM each) | Reconstruct the E. coli cytoplasm. K-glutamate over K-chloride because chloride inhibits many enzymes (Jewett & Swartz 2004). |
| Energy + nucleotides | Glucose, ribose, AMP, CMP, UMP, guanine — but no GMP | Drive NTP regeneration. See “Two batteries” below. |
| Building blocks | 17-AA mix + tyrosine (separate, low solubility) + cysteine (separate, oxidizes) | The amino acids. |
| Additive | Nicotinamide | Precursor for NAD⁺, which GAPDH needs to keep glycolysis running. |
| Backfill | Nuclease-free water | Volume only — no RNases allowed. |
Two batteries — the energy systems
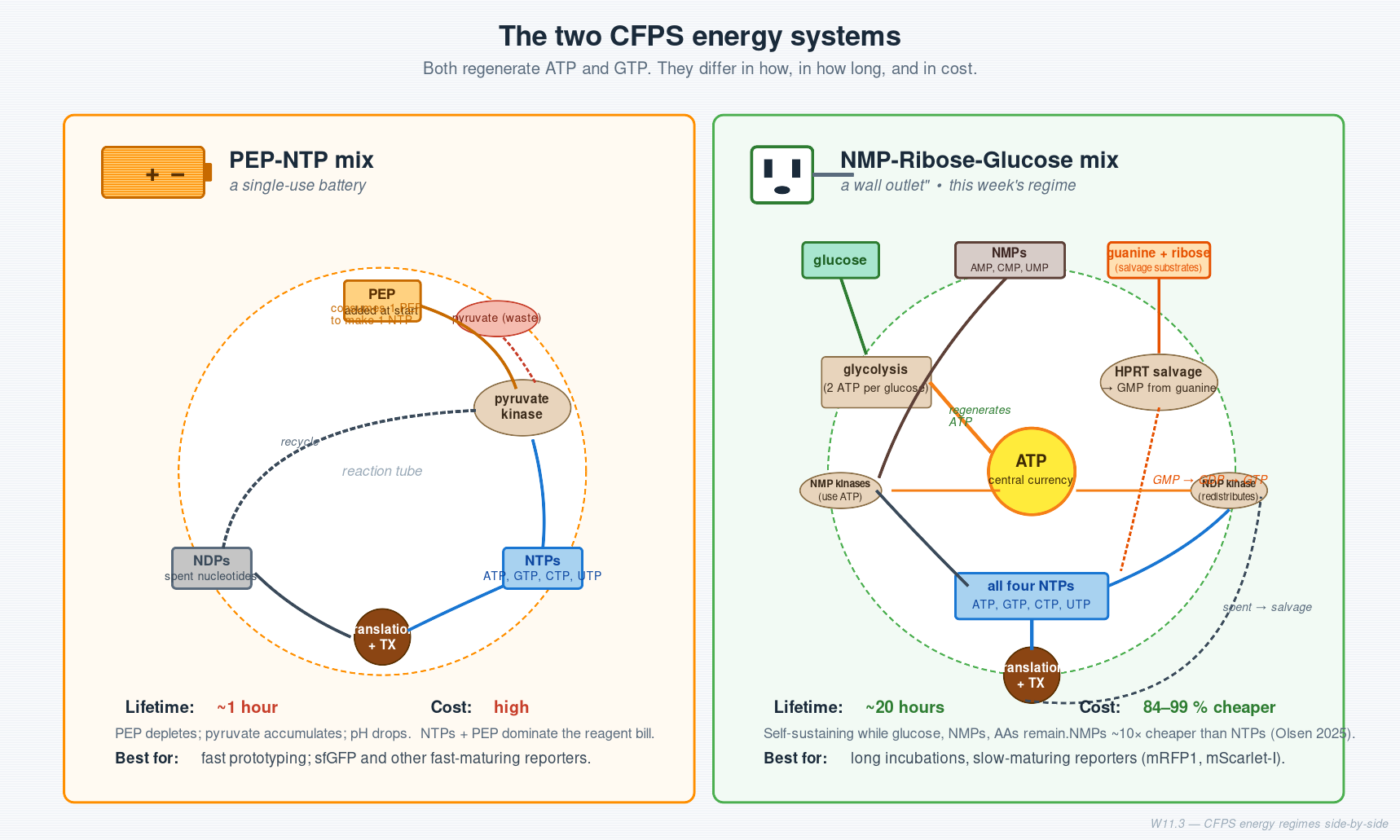
A CFPS reaction needs ATP and GTP regenerated continuously. There are two ways to do this:
| PEP-NTP mix | NMP-Ribose-Glucose mix (this week) | |
|---|---|---|
| Analogy | A single-use lithium battery | A wall outlet with a power adapter |
| What you add | Pre-made NTPs + PEP (high-energy phosphate donor) | Cheap NMPs + glucose + ribose + free guanine |
| How it regenerates | Pyruvate kinase puts PEP’s phosphate onto NDP → NTP | Glucose → glycolysis → ATP → NMP kinases + NDP kinase → all four NTPs |
| Runtime | ~1 hour | ~20 hours |
| Cost | High (NTPs are expensive) | 84–99 % cheaper (Olsen et al. 2025) |
| Best for | Fast prototyping, fast-maturing reporters | Long incubations, slow-maturing reporters, cost-sensitive work |
Worth the bold. The reason this week uses NMPs, glucose, and ribose instead of NTPs and PEP is that the cheaper system lets the reaction run 20× longer at roughly 1⁄10 the cost. That single trade is what makes the 1,536-well global canvas affordable to run.
The salvage shortcut — why guanine alone is enough
The master mix this week supplies no GMP. Only free guanine. The reaction still produces RNA, which needs GTP. How?
flowchart LR
G[Guanine] -->|"+ PRPP (from ribose + ATP)"| GMP
GMP -->|guanylate kinase + ATP| GDP
GDP -->|NDP kinase + ATP| GTP
GTP -->|RNA polymerase| RNA[Transcribed mRNA]
style G fill:#ffe
style GTP fill:#9cfThe lysate kept the cell’s purine salvage machinery. The key enzyme is HPRT (hypoxanthine-guanine phosphoribosyltransferase) — it takes guanine and PRPP (from ribose + ATP) and makes GMP. From there the usual kinases phosphorylate it up to GTP. As long as ribose and ATP are around, guanine alone carries the GTP pool. Bonus answer for Part B Q3.
The fluorescent proteins — five-property cheat sheet
Before any FP-specific reasoning, the five properties that decide whether a fluorescent protein works in CFPS:
| # | Property | Why it matters for CFPS |
|---|---|---|
| 1 | Maturation half-time | How long after translation before the chromophore is fluorescent. The dominant property for long reactions. |
| 2 | Brightness = EC × QY | Sets the detection floor. |
| 3 | pKa | CFPS metabolism acidifies the well. High pKa → signal fades as the reaction ages. |
| 4 | Oxygen dependence | All GFP/DsRed-family chromophores need O₂ to mature. Sealed wells run out within hours. |
| 5 | Oligomeric state | Monomers behave best. Dimers can self-quench at high local concentration. |
Photostability matters for imaging, not for endpoint reads.
The six FPs in this week’s canvas
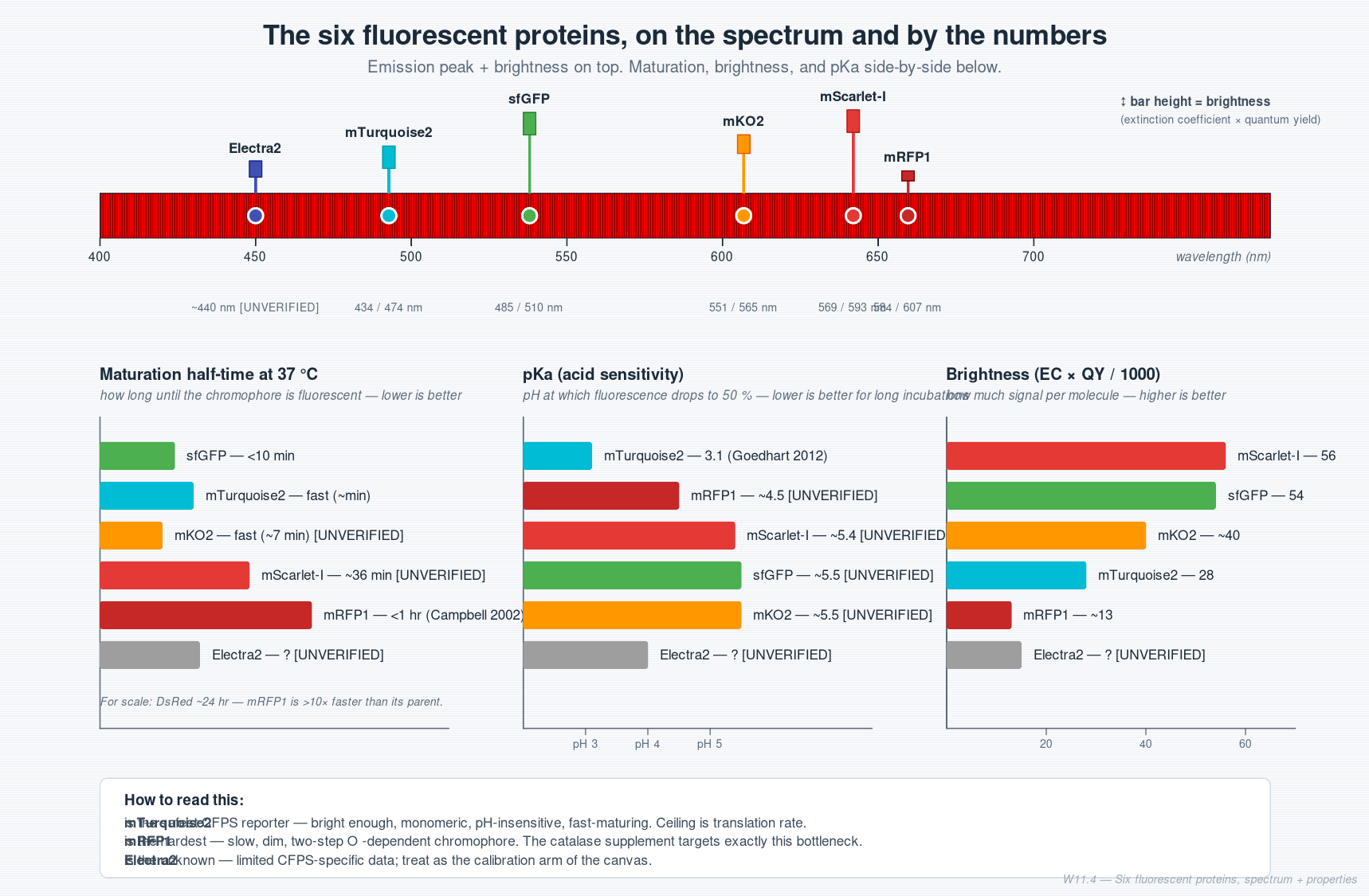
Each is chosen to expose a different bottleneck. The “supplement” column shows the per-well 2 µL intervention the dominant bottleneck argues for:
| FP | Dominant CFPS property | Suggested 2 µL supplement | Primary reference |
|---|---|---|---|
| sfGFP | Fast maturation (<10 min, 37 °C). No bottleneck. | Water control (no supplement needed). | Pédelacq et al. 2006, doi:10.1038/nbt1172 |
| mRFP1 | Slow, O₂-dependent two-step maturation (<1 hr, 37 °C — Campbell 2002 — vs DsRed’s ~24 hr). | Catalase, 100 U/mL — clears H₂O₂, recycles O₂. (See worked example.) | Campbell et al. 2002, doi:10.1073/pnas.082243699 |
| mKO2 | Moderate pKa (~5.5, per FPbase). Acid-sensitive at long incubation. | Boost K-phosphate to 15 mM + HEPES to 75 mM final. | Sakaue-Sawano et al. 2008, doi:10.1016/j.cell.2007.12.033 |
| mTurquoise2 | Highest QY of any monomer (0.93), pKa 3.1 — essentially pH-insensitive. Fast-maturing (Goedhart 2012 — the I146F structural mutation also accelerates folding); ceiling is translation rate. | Boost Mg²⁺ to 12 mM final. | Goedhart et al. 2012, doi:10.1038/ncomms1738 |
| mScarlet-I | T74I mutation = fast maturation. QY drops 0.70 (parent mScarlet) → 0.54 (mScarlet-I). EC ≈ 100,300 M⁻¹cm⁻¹ for the parent and largely preserved in mScarlet-I (the T74I substitution affects QY and folding kinetics, not the absorptive cross-section). Still a two-step chromophore. | Catalase, 100 U/mL (same as mRFP1; smaller effect expected). | Bindels et al. 2016, doi:10.1038/nmeth.4074 |
| Electra2 | Newer scaffold (2022, from Entacmaea quadricolor), limited CFPS-specific data. | Water control — treat as calibration arm of the canvas. | Papadaki et al. 2022, doi:10.1038/s41598-022-13214-0 |
Worked example — designing the mRFP1 well
mRFP1 is the most useful teaching case because its bottleneck — slow, O₂-dependent chromophore maturation — has a clean reagent fix.
How the mRFP1 chromophore actually matures
flowchart LR
A["Folded protein<br/>(Met-Tyr-Gly chromophore precursor)"] -->|"cyclize<br/>~1 min"| B["Cyclized intermediate<br/>(not fluorescent)"]
B -->|"+ O₂ → H₂O₂<br/>1:1 stoichiometry<br/>(Strack 2010)"| C["GFP-class green intermediate"]
C -->|"+ O₂ consumed<br/>(acylimine formation;<br/>H₂O₂ release inferred,<br/>not directly quantified)"| D["Mature red mRFP1<br/>584 / 607 nm"]
style D fill:#f88
style C fill:#9f9Two things go wrong in a sealed, 36-hour well:
- O₂ runs out. Starts at ~250 µM, depletes to single digits within hours. mRFP1 maturation stalls at the green intermediate.
- H₂O₂ builds up. Each oxidation step releases peroxide. Peroxide oxidizes the mature chromophore back to dark species, and oxidizes methionine/cysteine in the lysate machinery.
The fix
Add bovine liver catalase to the 2 µL supplement slot, 100 U/mL final. Catalase runs the reaction 2 H₂O₂ → 2 H₂O + O₂, which:
- clears the damaging peroxide, and
- regenerates one O₂ for every two peroxides consumed.
Both bottlenecks get hit by the same enzyme.
E. coli has its own catalases (KatE and KatG) and these are present in the lysate — but at limited concentration. Supplemental bovine liver catalase (Sigma C9322, the standard commercial source) boosts activity well above the lysate’s residual level, which is the rationale for adding it rather than relying on the native enzymes.
The 2× master mix (shared across the canvas)
| Component | 2× stock | Final (1×) | Why |
|---|---|---|---|
| HEPES-KOH pH 7.5 | 100 mM | 50 mM | Cytoplasmic-pH buffer. |
| K-glutamate | 260 mM | 130 mM | Ionic strength; glutamate avoids chloride enzyme inhibition. |
| Mg-glutamate (Mg²⁺) | 20 mM | 10 mM | Centered in the safe window. |
| K-phosphate dibasic | 20 mM | 10 mM | Phosphate pool + buffer. |
| K-phosphate monobasic | 20 mM | 10 mM | Buffer pair. |
| Glucose | 80 mM | 40 mM | Bumped above canonical 30 mM for the 36-hour endpoint. Reasoned extrapolation — Olsen et al. 2025’s reported optimum should be cross-checked before submission; if their value differs significantly, defer to theirs. |
| Ribose | 60 mM | 30 mM | PRPP precursor + pentose-phosphate flux. |
| AMP | 3 mM | 1.5 mM | NMP load. |
| CMP | 1.8 mM | 0.9 mM | NMP load. |
| GMP | 0 mM | 0 mM | Omitted by design — guanine + HPRT replaces it. |
| UMP | 1.8 mM | 0.9 mM | NMP load. |
| Guanine | 1 mM | 0.5 mM | Salvage substrate. |
| 17 AA mix | 3 mM each | 1.5 mM each | Building blocks. |
| Tyrosine | 3 mM | 1.5 mM | Separate (low solubility). |
| Cysteine | 3 mM | 1.5 mM | Separate (oxidizes); add fresh. |
| Nicotinamide | 2 mM | 1 mM | NAD⁺ salvage precursor. |
| NF-water | q.s. | — | Backfill. |
Three deliberate departures from canonical 20-hr NMP-RG: glucose elevated for the 36-hr endpoint; GMP fully omitted; Mg²⁺ centered rather than pushed high.
The 2 µL supplement (mRFP1-specific)
| Component | Stock | Final (in 20 µL) | Purpose |
|---|---|---|---|
| Bovine liver catalase | 1000 U/mL | 100 U/mL | H₂O₂ clearance + O₂ recycling. |
Expected outcome
Higher endpoint fluorescence than a no-supplement control. Magnitude unknown — there’s no direct DsRed-family + catalase CFPS literature precedent, so any specific multiplier would be speculation. The time-course should rise for longer, with no plateau-then-droop.
What we don’t know yet. — Whether the per-well 2 µL slot accepts enzyme supplements (confirm with the TAs). — Whether catalase keeps full activity in CFPS buffer (it should, but unverified). — Whether the second mRFP1 oxidation releases peroxide at exactly 1:1 (mechanistically required, less explicitly measured than the first step).
The bigger picture — the GPT-5 case study
The same experimental loop you’re running by hand was run by GPT-5 driving Nebula in late 2025, on the same kind of reaction:
flowchart LR
A[GPT-5 designs<br/>~256 conditions] -->|Pydantic<br/>validator| B[~128 feasible<br/>conditions]
B -->|"384-well plate<br/>78 conditions × 4 reps"| C[Nebula execution]
C -->|"titer + run metadata<br/>+ control QC"| D[GPT-5 lab notebook<br/>+ next-round hypotheses]
D -->|next round| A
style A fill:#cdf
style D fill:#cdf
style C fill:#fc9Six rounds, 36,000 conditions, 40 % cost reduction, 27 % titer increase vs the Olsen et al. 2025 SOTA. The most striking detail: in early rounds, before GPT-5 had access to the Olsen preprint, it independently proposed swapping NTPs for NMPs — the same insight that drove the prior SOTA.
This week’s homework deliberately puts you in the GPT-5 role — you do, by hand and reasoning, the parameter optimization Reshma argues should ultimately be outsourced to models. The point is to build the intuition for what that role actually requires.
Reshma’s framing of human × AI division of labor. Outsource parameter optimization (DOE, “which ten permutations next”) to the model. Humans pick the destination — the problem worth working on, the question worth asking. Waymo analogy: the human says where; the system handles speed, lane, turn.
The ethics tension, openly. A student raised that friction is where expertise lives — practical difficulty is a biosafety entry barrier. Reshma agreed: Ginkgo applies human review at the end-to-end research-services tier (won’t build bioweapons). The cloud-lab tier is currently data-in / data-out (CFPS, enzyme assays) and innocuous, so biosafety filters haven’t been forced. As the catalog expands and primitives can be chained, that conversation has to happen. The 50-state Cloud Lab Act legislation moving through Congress will force convergence.
Pitfalls & controls — how to know it worked
Run these alongside the optimized well:
| Control | Purpose |
|---|---|
| No-supplement (water in the 2 µL slot) | Baseline anchor. |
| Heat-inactivated catalase (boil stock 10 min at ≥95 °C, then add 2 µL as in the active condition) | Heat denatures the enzyme while preserving protein concentration in the well. If the active condition outperforms this, the gain is enzymatic — not generic protein-additive (osmotic, crowding) effects. |
| No-DNA (water in the DNA slot) | Lysate background fluorescence; subtract. |
| sfGFP positive control | Confirms lysate + master mix are competent. |
Common pitfalls:
- Sealing too tightly. Cuts off O₂; mRFP1 stalls at the green intermediate. Use a breathable film.
- Old cysteine. Cysteine that sat at 4 °C for >1 week is mostly oxidized cystine — replace.
- Mg²⁺ too high. Above ~15 mM, phosphate precipitates. Visible flocculent = remake.
- Endpoint-only reads. A plateau-then-droop diagnoses pH or peroxide problems; an endpoint number hides it. Request kinetic reads where possible.
Recommended reading
Four papers that anchor everything on this page:
- Olsen et al. 2025. Design-driven optimization of low-cost reagent formulations for reproducible and high-yielding cell-free gene expression. bioRxiv 2025.08.01.668204. doi:10.1101/2025.08.01.668204 — The NMP-Ribose-Glucose paper; 84–99 % cost reduction over PEP-NTP.
- Ginkgo Bioworks × OpenAI. 2026. Using a GPT-5-driven autonomous lab to optimize the cost and titer of cell-free protein synthesis. bioRxiv 2026.02.05.703998 — The 36,000-condition autonomous CFPS study; 40 % cost + 27 % titer beyond Olsen.
- Jewett & Swartz 2004. Mimicking the Escherichia coli cytoplasmic environment activates long-lived and efficient cell-free protein synthesis. Biotechnol Bioeng 86:19–26. doi:10.1002/bit.20026 — Foundational CFPS reagent paper; the K-glutamate-over-chloride paper.
- Goedhart et al. 2012. Structure-guided evolution of cyan fluorescent proteins towards a quantum yield of 93 %. Nat Commun 3:751. doi:10.1038/ncomms1738 — mTurquoise2 design; reference example for how structure-guided FP engineering pushes limits.
Course resources
- HTGAA Week 11: 2026a.htgaa.org/2026a/course-pages/weeks/week-11/
- 1,536-pixel bioart canvas: rcdonovan.com/1536
- Master-mix design tool: rcdonovan.com/cfps
- RACs simulator (Part D, optional): racs.rcdonovan.com
- FPbase entries: sfGFP · mRFP1 · mKO2 · mTurquoise2 · mScarlet-I · Electra2
- Nebula
Generic_*protocol modules referenced in the assignment:Generic_atc_run_protocol,Generic_bravo_stamp,Generic_cytomat_incubate,Generic_echo_hitpick,Generic_hig_centrifuge,Generic_floi8_cherry_pick,generic_multiflo_dispense,generic_spark_read,generic_pherastar_read.
References
- Olsen ML et al. 2025. bioRxiv 2025.08.01.668204. doi:10.1101/2025.08.01.668204.
- Ginkgo Bioworks × OpenAI. 2026. bioRxiv 2026.02.05.703998.
- Jewett MC, Swartz JR. 2004. Biotechnol Bioeng 86:19–26. doi:10.1002/bit.20026.
- Pédelacq JD et al. 2006. Nat Biotechnol 24:79–88. doi:10.1038/nbt1172.
- Campbell RE et al. 2002. PNAS 99:7877–7882. doi:10.1073/pnas.082243699.
- Sakaue-Sawano A et al. 2008. Cell 132:487–498. doi:10.1016/j.cell.2007.12.033.
- Goedhart J et al. 2012. Nat Commun 3:751. doi:10.1038/ncomms1738.
- Bindels DS et al. 2016. Nat Methods 14:53–56. doi:10.1038/nmeth.4074.
- Papadaki S et al. 2022. Sci Rep 12:10190. doi:10.1038/s41598-022-13214-0.
- Strack RL et al. 2010. J Am Chem Soc 132:8496–8505. doi:10.1021/ja1020364.