Individual Final Project



Flavonoids are abundant and widely distributed plant secondary metabolites. They are the primary compounds of plant pigments, provide signals for pollinators and symbiotic bacteria, protect plants from UV-B and environmentally induced oxidative stress. 3
The ability to manipulate plant pigmentation has been extensively studied, particularly in ornamental flowers such as tulips or petunia. Flower color is primarily determined by anthocyanins, a class of flavonoid pigments responsible for red, purple and blue hues. In species like tulipa, variation in color is achieved through differences in anthocyanin composition, concentration and cellular localization. Genetic engineering approaches have successfully modified flower color by altering key enzymes in the anthocyanin biosynthesis pathway or by introducing transcription factors that regulate pigment production. 4
For example, previous studies have demonstrated that overexpression or suppression of enzymes such as dihydroflavonol reductase (DFR) or anthocyanidin synthase (ANS) can shift pigmentation outcomes. Additionally, regulatory genes such as MYB transcription factors have been used to activate entire pigment pathways, enabling predictable and stable color changes in petals. These advances illustrate that plant pigmentation can be rationally engineered when the biosynthetic pathway and its regulation are well understood. 5
In contrast to ornamental flowers, pigmentation in fiber-producing crops such as Gossypium hirsutum remains largely unexplored as a designable trait. Cotton fibers are single elongated epidermal cells composed primarily of cellulose and are typically white or off-white. While naturally colored cotton varieties exist (e.g. brown or green), their color range is limited and not easily tunable. 6
However, existing research shows that cotton is capable of producing anthocyanins under specific conditions. In response to infection by the bacterial pathogen Xanthomonas campestris pv. malvacearum, cotton accumulates red pigmentation at infection sites. This coloration is caused by anthocyanins, particularly cyanidin-3-glucoside, which play a protective role by absorbing light and mitigating damage from reactive oxygen species and light-activated phytoalexins. Importantly, studies have shown that these pigmented cells can absorb 3–4 times more photo-activating light, protecting surrounding healthy tissue. 78
This demonstrates that:
- Cotton possesses a functional anthocyanin biosynthesis pathway
- Pigment production is inducible and spatially regulated
- Pigmentation is linked to stress and defense responses, not material development
Knowledge Gap
This project addresses this gap by proposing to reprogram the regulatory control of anthocyanin biosynthesis in cotton, shifting it from a stress-induced response to a developmentally controlled trait.
By drawing on established strategies from flower color engineering (e.g. transcription factor activation and pathway modification) and applying them to cotton, this work explores the possibility of creating intrinsically colored fibers. This represents a novel intersection of synthetic biology, plant science and material design, with potential applications in sustainable textile production.
Despite advances in engineering pigmentation in flowers, there is a clear gap in applying these strategies to fiber-producing plant tissues such as cotton. Specifically:
- Anthocyanin production in cotton is restricted to stress conditions and does not occur during normal fiber development
- There is currently no established method to program pigment production directly into cotton fibers
- The integration of pigmentation into fibers during growth remains unexplored 9
Innovation
This project could be innovative in that it proposes a shift from post-production dyeing to biologically embedded color, reframing pigmentation as a material property that is designed during growth rather than applied afterward. While anthocyanin pathways have been extensively engineered in ornamental plants for aesthetic purposes, their application to fiber-producing crops like Gossypium hirsutum remains largely unexplored. By transferring and adapting these biological concepts, the project introduces a new use of existing genetic tools to address environmental challenges in the textile industry.Furthermore, the work challenges the prevailing assumption that pollution must be managed through downstream solutions such as filtration, instead proposing a preventative, design-based approach rooted in synthetic biology. It expands the boundaries of the field by positioning plants not only as organisms to be engineered for yield or resistance, but as programmable material systems capable of producing functional and aesthetic properties simultaneously.
Summary of two peer-reviewed research citations
Chandler, S., Tanaka Y. (2007) Genetic Modification in Floriculture, Critical Reviews in Plant Sciences, 26:4, 169-197, https://doi.org/10.1080/07352680701429381 4
Research by Stephen Chandler and Yoshikazu Tanaka (2007) provides a comprehensive review of genetic modification in floriculture, focusing particularly on the engineering of flower color. The study explains that traditional breeding is limited by the natural gene pool of a species, whereas genetic modification enables the introduction of new genes to create novel traits, especially through manipulation of the anthocyanin biosynthesis pathway. A key achievement highlighted is the development of genetically modified carnations with new colors (e.g. violet/blue hues), demonstrating that pigment pathways can be successfully reprogrammed to produce traits not naturally present in the plant. However, the authors note that despite significant scientific progress, commercial applications remain limited due to regulatory costs, intellectual property constraints, and perceived public acceptance issues.
Shi, S.; Tang, R.; Hao, X.; Tang, S.; Chen, W.; Jiang, C.; Long, M.; Chen, K.; Hu, X.; Xie, Q.; et al. (2024) Integrative Transcriptomic and Metabolic Analyses Reveal That Flavonoid Biosynthesis Is the Key Pathway Regulating Pigment Deposition in Naturally Brown Cotton Fibers. Plants, 13, 2028. https:// doi.org/10.3390/plants13152028 8
Research on pigmentation in Gossypium hirsutum demonstrates that naturally colored cotton fibers derive their pigmentation primarily from the flavonoid biosynthesis pathway. A recent transcriptomic and metabolomic study (Shi et al., 2024) showed that key genes such as CHS, DFR, F3H, and UFGT are significantly upregulated in brown cotton fibers, particularly during later developmental stages. The study also identified metabolites including cyanidin-3-O-glucoside as major contributors to fiber coloration, and highlighted the role of MYB transcription factors in regulating pigment production. However, despite identifying these pathways and regulatory networks, the study concludes that the mechanism of pigment deposition in fibers remains poorly understood, limiting the ability to engineer new or controllable colors in cotton.
In contrast, research on ornamental plants such as Tulipa has demonstrated that anthocyanin-based pigmentation can be precisely engineered through genetic modification. Studies have shown that altering the expression of biosynthetic enzymes (e.g. DFR, ANS) or regulatory transcription factors (e.g. MYB proteins) enables predictable changes in flower color. These systems illustrate that pigmentation pathways can be externally controlled and fine-tuned, resulting in a wide spectrum of stable colors. Together, these studies highlight a key gap: while pigmentation in flowers is highly programmable, cotton fibers possess similar biochemical pathways but lack the regulatory control needed for designed and scalable color production.
Why this project matters?
It addresses a fundamental limitation in the textile industry: the reliance on post-production dyeing, which leads to significant environmental pollution. In my Bachelor thesis, I conducted a life cycle assessment of a cotton T-shirt grown in India, manufactured in Bangladesh, and sold in Austria. The results challenged common assumptions: contrary to expectations, transportation contributed only minimally to the overall impact, accounting for approximately 0.1 kg CO₂-eq, whereas the production phase—particularly textile finishing — was responsible for nearly 4 kg CO₂-eq for a 200 g cotton T-shirt. Additionally, I found that dyeing alone can require around 5 liters of water per T-shirt, highlighting the disproportionate environmental burden of this stage. These findings emphasize that addressing the finishing process is critical for achieving meaningful reductions in the environmental footprint of clothing. The wastewater generated from textile dyeing presents substantial environmental and social challenges, it threatens access to safe water for drinking, sanitation and hygiene, while also damaging aquatic ecosystems. (SDG Goals 6 & 14) Common dye classes—including naphthalene-based, heterocyclic, anthraquinone, and indigo dyes—are associated with serious health risks such as skin irritation, respiratory issues, carcinogenic effects, and liver and kidney damage. On a global scale, it is estimated that approximately 280,000 tons of textile dyes are discharged into wastewater annually, with up to 80% released untreated into the environment, exacerbating ecological and human health impacts. As highlighted in recent research on Gossypium hirsutum, pigment production in cotton fibers is already biologically possible through the flavonoid biosynthesis pathway. However, this mechanism is not yet controllable or scalable for industrial use, representing a key barrier to progress. While naturally colored cotton offers a promising alternative, its limited color range, instability and inconsistent expression prevent widespread adoption and maintain dependence on chemical dyeing. 10
Building on this gap, the project proposes to move beyond observation toward engineering controllable pigmentation directly within the fiber, eliminating the need for external dyes. The potential impact is significant: reducing water consumption, chemical inputs and energy use across the textile lifecycle. Beyond environmental benefits, this approach could fundamentally transform how materials are designed, enabling fibers to possess both functional and aesthetic properties from the moment they are grown.
Bioethics & Biosafety
This project raises several ethical considerations related to the use of genetic modification in agriculture, particularly in a widely cultivated species such as Gossypium hirsutum. The deliberate alteration of plant metabolic pathways to produce colored fibers introduces questions about biosafety, ecological impact and long-term consequences. For example, engineered pigment pathways could unintentionally affect plant fitness, interactions with pests or surrounding ecosystems if gene flow occurs. Working with GMOs requires careful design, testing and containment strategies to be certain that innovations in synthetic biology are applied safely. In addition, the cultivation of cotton - as one of the most relevant crops - engages with broader ethical questions around justice and accessibility. Textile pollution disproportionately affects communities in major production regions, where untreated wastewater impacts local water resources and public health. By proposing a preventative alternative, this work has the potential to contribute to a more equitable distribution of environmental burdens. However, it is also important to consider who benefits from such innovations: access to genetically modified seeds, intellectual property rights and economic implications for hydroponic farmers must be addressed to avoid reinforcing existing inequalities. Transparent communication, inclusive decision-making and consideration of local contexts are therefore essential to ensure that the benefits of this technology are shared fairly, aligning the project with principles of justice and global responsibility.
Measures
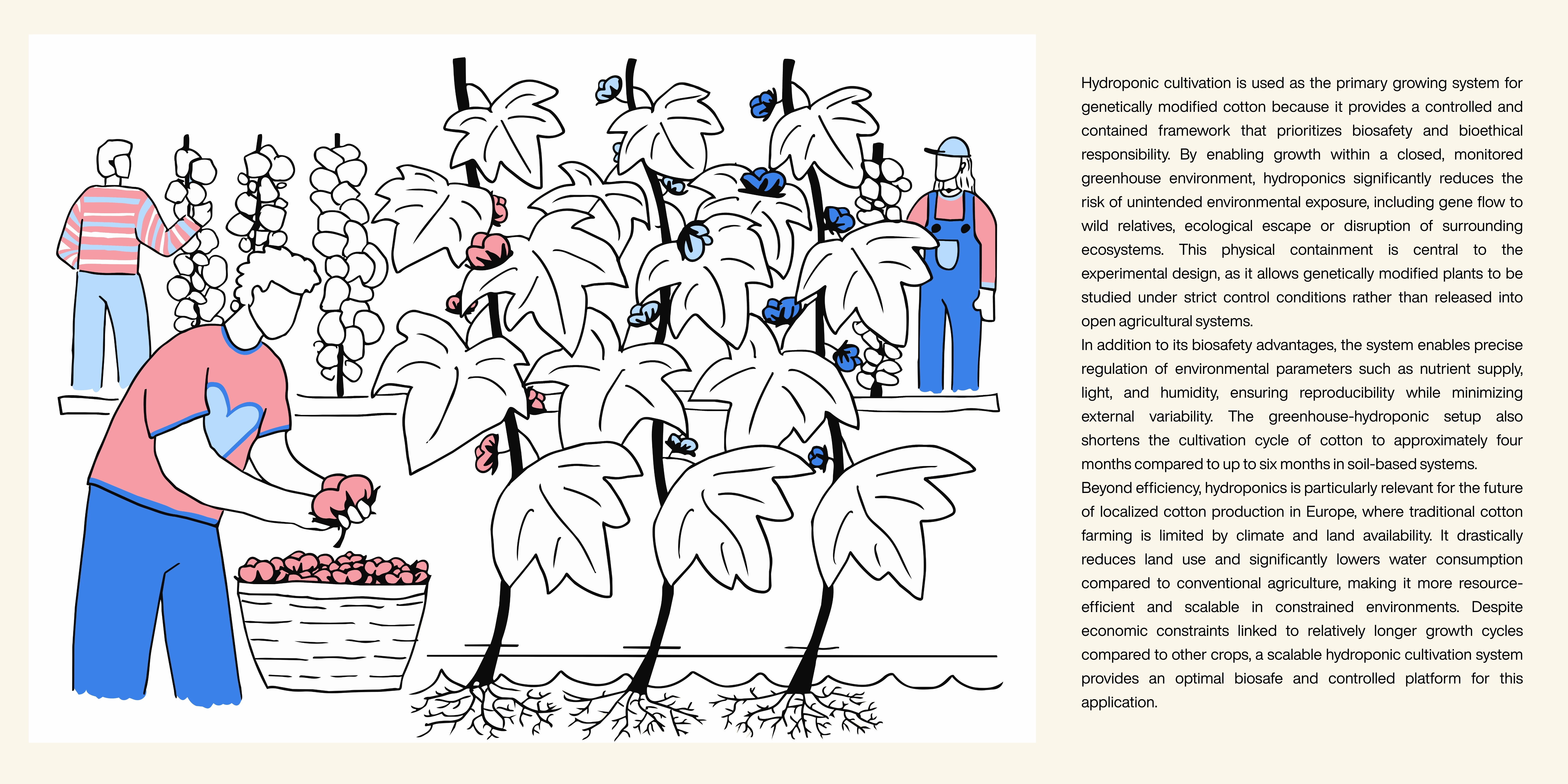
To ensure that this project is conducted ethically, it will be implemented exclusively within hydroponic systems in closed, monitored greenhouse environments. Restricting cultivation to controlled settings significantly reduces the risk of unintended environmental exposure, including gene flow to wild relatives, ecological escape, or disruption of surrounding ecosystems. Nevertheless, even when plants are grown hydroponically without contact with soil, it remains essential to conduct controlled greenhouse trials, apply gene-flow mitigation strategies, and perform comprehensive ecological risk assessments. Continuous monitoring is necessary to evaluate potential unintended consequences, such as impacts on plant metabolism, ecosystem interactions, or the spread of modified traits beyond the intended system. Transparency and open communication with stakeholders — including farmers, policymakers, and the public — should be integrated throughout the research process to foster trust and support informed decision-making. Potential unintended consequences may include unforeseen ecological effects, limitations in color diversity or durability, and socioeconomic challenges such as unequal access to modified seeds or dependency on proprietary technologies. These uncertainties underline the importance of iterative testing and interdisciplinary collaboration. To put it in a nutshell, colorful grown cotton aims to contribute to society by indirectly improving equitable access to safe drinking water, reducing potential health risks for workers in conventional dyeing facilities, and relocating cultivation closer to the areas where the products are sold, thereby potentially decreasing transport-related environmental impacts.

I will use databases and Benchling to organize and develop the genetic constructs needed for engineering naturally colorful cotton. Scientific databases such as NCBI GenBank and UniProt would help me research pigment-producing genes, regulatory elements and existing studies related to plant coloration and cotton genetics. In Benchling, I will design my expression casette, annotate it and export it to Twist Bioscience.
Additionally, designing a Twist order will be an important step in transforming the digital genetic designs into physical DNA constructs. Through Twist Bioscience, I would order synthesized DNA fragments containing the selected pigment genes and promoters optimized for cotton expression. This process would allow me to efficiently move from computational design to experimental implementation in developing naturally colorful cotton fibers.
Experimental plan (7-9 weeks)
1. Literature Research into flavonoid pathway Literature Research on the Flavonoid Biosynthesis Pathway (1–2 weeks) The project will begin with an extensive literature review on the flavonoid biosynthesis pathway, with a particular focus on anthocyanin and pigment production in plants. Scientific databases such as Google Scholar and NCBI will be used to identify key enzymes, regulatory genes, and previous genetic engineering approaches related to pigmentation in plants. Expected result: Identification of the most relevant enzymes and pathways involved in stable pigment formation.
2. Selection of and Specialized Research into a Gene of Interest (1 week) After comparing multiple candidate genes, one gene involved in anthocyanin biosynthesis will be selected based on criteria such as pigment intensity, stability, previous use in plant engineering studies, and compatibility with cotton. Particular attention will be given to genes reported to strongly influence visible coloration. Once a candidate gene has been chosen, a more detailed literature review will investigate its biological function, regulation, enzymatic activity, and performance in previous transformation experiments. Expected result: Selection of the most promising pigmentation-related gene and further investigation.
3. Retrieval of Gene Sequences from Multiple Species (3–4 days) DNA and amino acid sequences of the selected gene will be retrieved from databases such as NCBI GenBank and UniProt. Sequences from multiple plant species will be collected to compare natural variation and identify the most suitable version for expression in cotton. Expected result: A collection of candidate sequences from different species for comparative analysis.
4. Reverse Translation of Amino Acid Sequences and Codon Optimization for Gossypium hirsutum (2 days) Amino acid sequences of the selected protein will be reverse translated into nucleotide sequences using the Reverse Translate Tool from Bioinformatics.org, subsequently the nucleotide sequence will be codon optimized for cotton using the VectorBuilder Codon Optimization Tool to improve predicted protein expression efficiency. Parameters such as GC content and codon adaptation index will be evaluated. Expected result: An optimized gene sequence predicted to express efficiently in cotton cells.
5. Evaluation of Expression Efficiency (3–4 days) Computational analysis will compare optimized and non-optimized sequences to assess possible differences in translational efficiency, sequence stability, and mRNA structure. Expected result: Evidence that codon optimization improves theoretical expression potential.
6. Research into Regulatory DNA Elements (1 week) Suitable promoters, terminators, and regulatory elements for plant transformation will be identified through literature research. Expected result: Selection of appropriate regulatory sequences for stable gene expression.
7. Design of the Expression Cassette (3–5 days) An expression cassette containing the promoter, RBS, start codon, optimized gene sequence, his tag, terminator and stop codon will be digitally assembled. Expected result: A complete DNA construct for hypothetical plant transformation.
8. Selection, Assembly and Validation of the Plasmid Construct (4–6 days) An appropriate plasmid vector for plant transformation will be selected based on cloning compatibility, selectable markers and compatibility with Agrobacterium tumefaciens systems. The designed expression cassette will then be integrated into the selected vector and uploaded to Twist Bioscience to evaluate whether the sequence is technically synthesizable and free of problematic sequence characteristics such as repetitive regions or unstable motifs. Expected result: Identification of a suitable plasmid vector and confirmation that the construct is feasible for synthetic DNA production.
9. Visualization, Annotation, and Optimization of the Construct (1 day) The completed plasmid construct will be imported into Benchling to generate a detailed annotated plasmid map. Functional regions including promoters, coding sequences, terminators, and selectable markers will be clearly labeled. The construct will then be analyzed for unwanted restriction sites, sequence instabilities, or cloning incompatibilities, and refinements will be made where necessary to optimize the design for stable expression. Expected result: A fully annotated, refined and optimized plasmid construct suitable for hypothetical plant transformation experiments.
10. Creation of Visual Workflow and Plasmid Graphs (1 week) Diagrams and graphs will be produced to visualize the project workflow, plasmid composition, and sequence proportions. These figures will support communication of the experimental design. Expected result: Clear visual materials illustrating the genetic engineering workflow and construct design.
11. Final Analysis and Ethical Assessment (1 week) The completed project design will be evaluated regarding feasibility, sustainability, biosafety, and societal implications. Potential ecological risks, greenhouse containment strategies, and ethical considerations surrounding genetically modified crops will be discussed. Expected result: A comprehensive assessment showing how the project addresses both scientific feasibility and responsible innovation.
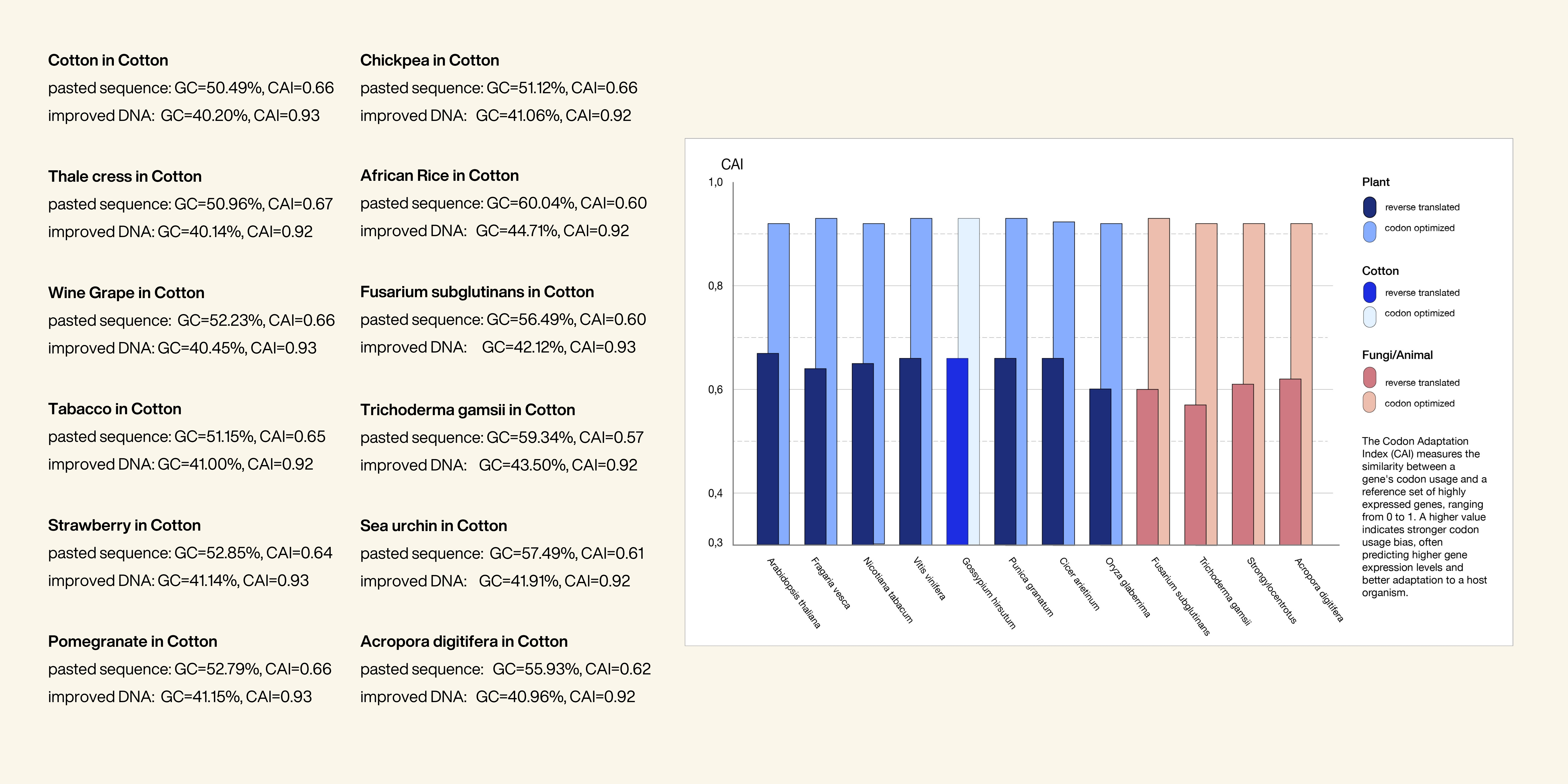
After extensive research, I decided to focus on Dihydroflavonol 4-reductase, or DFR. DFR is a key enzyme in the flavonoid biosynthetic pathway and plays an essential role in the production of anthocyanin pigments - the compounds responsible for red, purple, and blue coloration in plants. Beyond coloration, these pigments contribute to UV protection and can support plant stress responses. Instead of introducing an entirely foreign protein into the organism, I wanted to work with a naturally existing pathway and explore how its expression could be intensified. Therefore, I experimented with codon optimization of the DFR gene itself in order to potentially increase expression efficiency while maintaining biological compatibility. This approach reflects the idea that laboratory experimentation is necessary to evaluate feasibility, but in this case, optimizing a native system appeared to be the most plausible and sustainable strategy. As shown in the graphic, I compared DFR gene sequences from multiple plant species. First, I generated the nucleotide sequences using the Reverse Translate tool from Bioinformatics.org. Afterwards, the sequences were codon-optimized specifically for Gossypium hirsutum, or upland cotton, using the official VectorBuilder codon optimization tool. The optimization results did not reveal major differences between the variants. Because of this, I ultimately selected the cotton-derived DFR sequence, as it showed a relatively high Codon Adaptation Index, or CAI, value of 0.93 and offered the advantage of native compatibility with the host organism. But as a next step, and if I had access to a lab I would love for my aim 2 to experiment with DFR from multiple species - especially ones that have an intense red/purple/blue coloration, like strawberries or grapes. It seems there must be a difference because cotton has the same gene but is still not appearing red or blue. In the following points, you can see the individual steps of my working process and how I developed this experimental approach.
Fasta File Text
tr|C8YQV2|C8YQV2_GOSHI Dihydroflavonol 4-reductase OS=Gossypium hirsutum OX=3635 GN=LOC107957957 PE=2 SV=1 MGSSVTDGEIVCVTGGSGFIGSWLIKLLLERGYVVRATVRDPGNSKKVKHLLELPKAETHLTLWKADLAEEGSFDDAIQACTGVFHVATPMDFESEDPENEVIKPTINGVLSIMKACAKAKTVRRLVFTSSAGTIDVAEQQKPCYDETCWSDLEFIQAKKMTGWMYFVSKTMAEQAAWKFAKENNIDFVSIIPPLVVGPFIMQSMPPSLITALSPITGNEAHYSIIKQGQFIHLDDLCRAHIFLFENPKAEGRHICASHHATIIDLAKMLSEKYPEYNVPTKFKDVDENLKSVEFSSKKLLDLGFEFKYSLEDMFVGAVETCREKGLLPLSNEKKIKNID 13
AlphaFold 3D Protein Structure
Reverse Translate: Protein (amino acid) sequence to DNA (nucleotide) sequence
reverse translation of tr|C8YQV2|C8YQV2_GOSHI Dihydroflavonol 4-reductase OS=Gossypium hirsutum OX=3635 GN=LOC107957957 PE=2 SV=1 to a 1020 base sequence of most likely codons.
atgggcagcagcgtgaccgatggcgaaattgtgtgcgtgaccggcggcagcggctttattggcagctggctgattaaactgctgctggaacgcggctatgtggtgcgcgcgaccgtgcgcgatccgggcaacagcaaaaaagtgaaacatctgctggaactgccgaaagcggaaacccatctgaccctgtggaaagcggatctggcggaagaaggcagctttgatgatgcgattcaggcgtgcaccggcgtgtttcatgtggcgaccccgatggattttgaaagcgaagatccggaaaacgaagtgattaaaccgaccattaacggcgtgctgagcattatgaaagcgtgcgcgaaagcgaaaaccgtgcgccgcctggtgtttaccagcagcgcgggcaccattgatgtggcggaacagcagaaaccgtgctatgatgaaacctgctggagcgatctggaatttattcaggcgaaaaaaatgaccggctggatgtattttgtgagcaaaaccatggcggaacaggcggcgtggaaatttgcgaaagaaaacaacattgattttgtgagcattattccgccgctggtggtgggcccgtttattatgcagagcatgccgccgagcctgattaccgcgctgagcccgattaccggcaacgaagcgcattatagcattattaaacagggccagtttattcatctggatgatctgtgccgcgcgcatatttttctgtttgaaaacccgaaagcggaaggccgccatatttgcgcgagccatcatgcgaccattattgatctggcgaaaatgctgagcgaaaaatatccggaatataacgtgccgaccaaatttaaagatgtggatgaaaacctgaaaagcgtggaatttagcagcaaaaaactgctggatctgggctttgaatttaaatatagcctggaagatatgtttgtgggcgcggtggaaacctgccgcgaaaaaggcctgctgccgctgagcaacgaaaaaaaaattaaaaacattgat
By comparing the original and optimized sequences, I increased the codon adaptation index (CAI) from 0.66 to 0.93, meaning the redesigned DNA now uses codons that better match those found in highly expressed cotton genes. This was especially interesting since I learned that even native genes in the same organism often have suboptimal codon usage, regulatory constraints and evolutionary tradeoffs. This suggests the optimized sequence would likely be translated more efficiently, potentially increasing enzyme production in the flavonoid pathway. ATGGGTTCATCTGTTACAGATGGTGAGATTGTTTGTGTCACTGGAGGTTCTGGATTCATCGGATCATGGCTTATTAAACTTCTTCTTGAAAGGGGTTATGTTGTTAGAGCTACAGTTAGAGATCCAGGAAACTCAAAGAAGGTTAAACACCTTTTGGAGCTTCCTAAGGCCGAGACTCATCTCACCCTTTGGAAGGCCGATTTGGCTGAAGAGGGTTCATTTGATGATGCTATTCAAGCATGTACCGGTGTTTTTCATGTGGCTACTCCTATGGATTTCGAGTCTGAGGATCCTGAAAACGAGGTTATCAAGCCTACAATCAACGGTGTTTTGTCCATCATGAAGGCATGCGCTAAAGCTAAGACTGTTAGGAGATTGGTTTTCACTTCCTCAGCTGGTACTATCGATGTTGCTGAACAACAAAAGCCTTGCTACGATGAGACATGTTGGTCTGATTTGGAATTCATTCAAGCTAAGAAGATGACCGGATGGATGTATTTCGTTTCAAAGACAATGGCCGAACAAGCAGCTTGGAAGTTTGCTAAGGAAAACAATATAGATTTCGTTTCAATTATCCCACCACTTGTTGTAGGACCTTTCATTATGCAATCCATGCCTCCTTCTCTTATTACTGCTTTGAGTCCCATTACTGGTAATGAGGCTCATTATTCTATTATTAAACAAGGACAATTTATTCATCTTGATGATCTTTGTAGAGCCCATATTTTTTTGTTCGAAAACCCTAAAGCAGAGGGTAGGCATATTTGTGCTTCCCATCATGCTACTATTATCGATCTTGCTAAGATGCTTTCAGAGAAGTATCCTGAATATAACGTTCCTACTAAGTTCAAGGATGTCGATGAAAACTTGAAATCTGTTGAATTCTCCTCAAAGAAACTCTTGGATCTTGGATTTGAATTCAAGTACTCTCTTGAAGATATGTTCGTTGGGGCTGTTGAGACTTGCAGGGAGAAGGGACTTCTGCCTCTTTCAAACGAGAAGAAAATTAAGAACATTGAT
Promoter
The SCFP promoter fragment was extracted from the GenBank entry (GQ411495.1) of Gossypium hirsutum cultivar Handan 5833 fiber-specific protein (SCFP) gene, promoter region and 5’ UTR, which contains a 1005 bp sequence annotated as promoter region plus 5’ UTR/mRNA. The promoter region corresponds to nucleotides 1–633, while nucleotides 634–1005 correspond to the 5’ UTR/mRNA. 14 13 15 16
ACAACTTTTCTCTACCAATCAAATTTAAAAAATAGAAAAATGAAAATCGATGAATTGGATCACCACAATTTAGCCCAAAGAAAAACACAGTCAACCCCTCTCACAGGGTAGGAATGATTTCGAGGTATAGATAGACATAGTAACGGGCAACTTTAACTATTGCTGCCTCGATTTGAGGAAAATATCAAATCCAAGACAAAAATTTCAATTATACACTATGCCATACCATTATAAATATCCCCGTTCGCAATATCATCACCATTATTTGAATTTGCATTGCAACATTCGTCACCGTTAGTTATACCATCACCATCACTTAATTACTAAAATAATTATTGGTTTCTCAATATGAAAAAGCTCGAGTGCATTTTCTTTTGAATATCAACCGAAAAGAAAGGAAAAACTAAAGA TTTTGGAAGGTGACGGGGAAACCAAAAAGGAAATTTTGGGCATTTTTAAAATGAGAAAGACGAATGTAATAACCCATTTTTCTTTCTTACTCTGACAACGCCACAGATGCTTTACATGCATCATGTGATCGTGGGGGACCCGAAACTTGGCATACGGAAAGCACCAACGGCACAGCATTAAAAGAAATTGTGTATAATGTTAAAAGACCATTAATTCAGTCTC
No RBS but 5’ UTR sequence
Because cotton is a eukaryotic plant and RBSs only work in prokaryotes (bacteria). 17
ATCCAACCACGCTTAAAAGTCTTCATGCCTTTTCTCACCTCTGATTTCATCTAATGAAAAGCGGACAAGTTGAAGGATCACTCGTTGCTTGTGTGAGCTTTCATTATTTATTATTATGTTTTAGGTAACCATAGGAAGAAGCCATTAACAACAGCATGAAAAACAGCTAGTTTCTCCGCAAACAAGATAAACTTTTATACTTTTTACCACTGCACCCCCCCCCAAAGACCAGTTTTTAACTCCACCTACCAAGCATTCAAGAAGCACCAACCAACTTAATTACCAGCTTAACAAGACAGTACAGGTTTCTGGGATATTTGTAGTCTCTCAAGGACATCACCACCTCCACTCACCTTCCCATTTTTCTCTAGC
Start Codon
ATG
6x His tag
CACCATCACCACCATCAC
I chose a 6×His tag because it is a small and widely established affinity tag that enables efficient detection and purification of the recombinant protein without significantly increasing the size of the expressed sequence. Compared to longer variants such as 7×His, the 6×His tag is more commonly used in standard molecular biology workflows and generally provides sufficient binding affinity. In the context of DFR expression, using a small tag is advantageous because it minimizes the risk of interfering with protein folding or enzymatic activity.18
Stop codon
The TAA (UAA in mRNA) stop codon was selected for the cotton DFR construct because its usage pattern in plants suggests it is the most efficient and reliable termination signal, particularly in dicotyledonous species. In dicots, UAA is the most frequently used stop codon, occurring in approximately 46% of genes, whereas UGA is used in about 36% and UAG in only 18%. This distribution indicates a clear preference for UAA in dicot genomes, suggesting better compatibility with the endogenous translation termination machinery. In contrast, UAG is comparatively underrepresented and has been associated with stronger context dependence, which may increase the likelihood of inefficient termination or readthrough. Therefore, selecting UAA provides a biologically supported choice that aligns with natural codon usage bias in dicots and supports efficient translation termination in plant expression systems. 19
nos terminator
The NOS terminator was selected because it is a well-established and reliable regulatory sequence commonly used in plant genetic engineering. The terminator ensures proper transcription termination and supports consistent and stable expression of the introduced construct. Its compatibility with plant transformation systems and extensive use in previous studies also made it a suitable and low-risk choice for achieving stronger color development in the cotton fibers.
GATCGTTCAAACATTTGGCAATAAAGTTTCTTAAGATTGAATCCTGTTGCCGGTCTTGCGATGATTATCATATAATTTCTGTTGAATTACGTTAAGCATGTAATAATTAACATGTAATGCATGACGTTATTTATGAGATGGGTTTTTATGATTAGAGTCCCGCAATTATACATTTAATACGCGATAGAAAACAAAATATAGCGCGCAAACTAGGATAAATTATCGCGCGCGGTGTCATCTATGTTACTAGATC
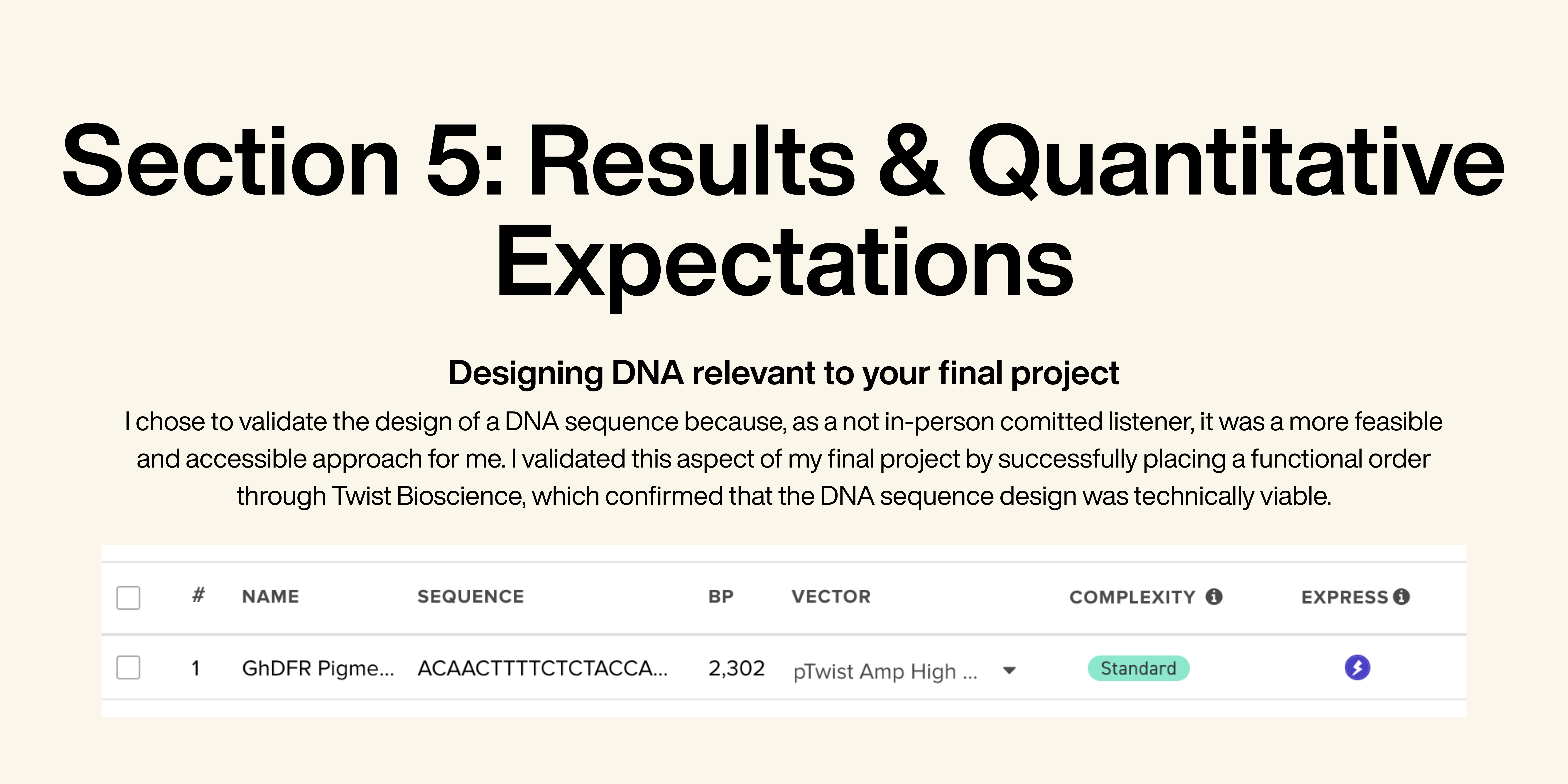
I imported all the DNA fragments into Benchling one after another and annotated the sequence for the first time. After exporting the annotated sequence as a FASTA file, I imported it into Twist Bioscience and selected the vector pTwist Amp High Copy. The order validation worked successfully, confirming that the construct was functional.
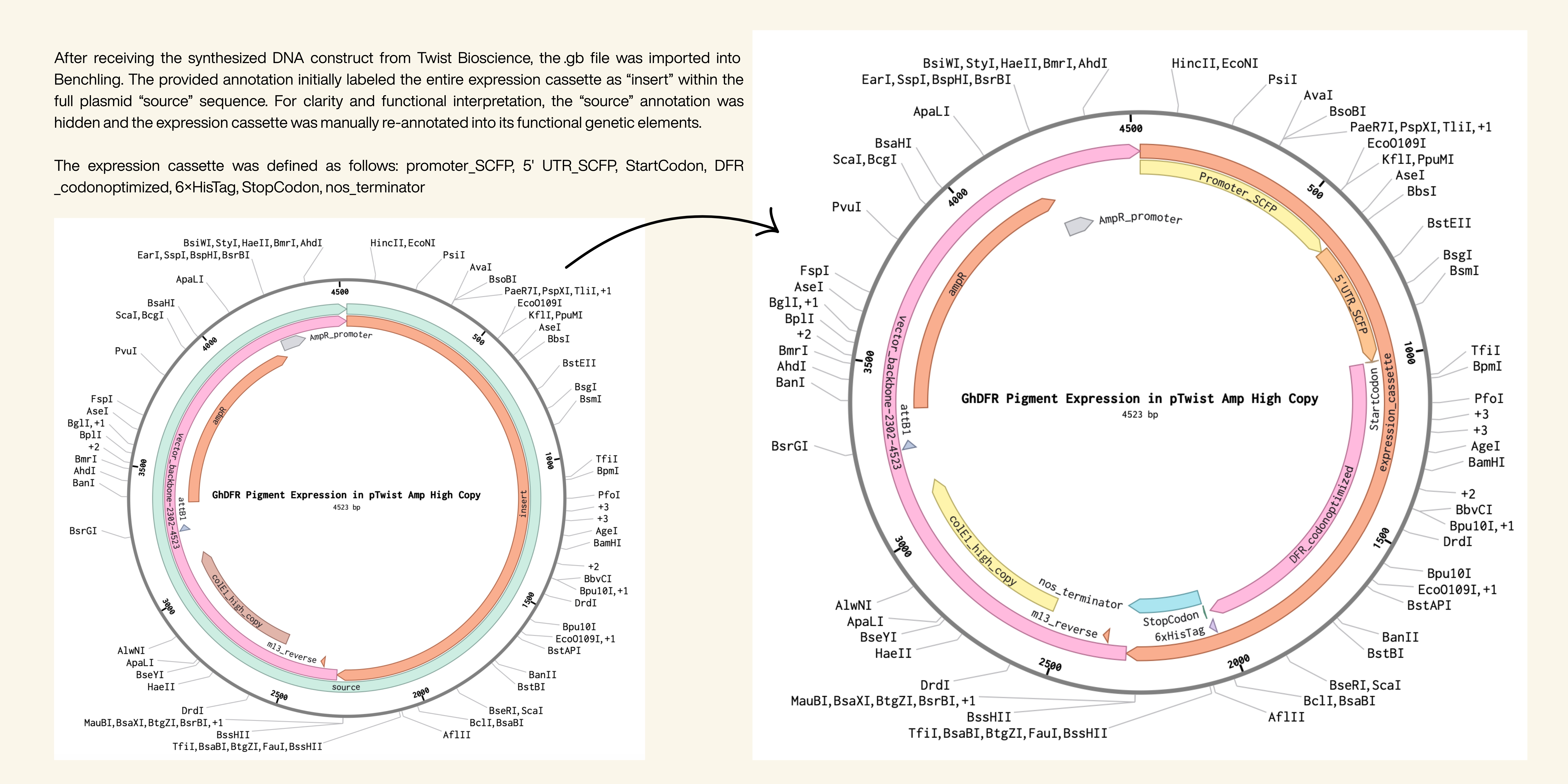
Key Synthetic Biology Techniques for Realization
I utilized several core synthetic biology techniques, including DNA sequencing, DNA construct design, databases, and designing a Twist order. DNA sequencing was important for analyzing and comparing DFR gene sequences from multiple plant species in order to identify suitable variants for anthocyanin production. I used publicly available genetic databases such as NCBI GenBank and protein information from UniProt to obtain the original sequences and study their biological function within the flavonoid biosynthesis pathway. Another important technique was DNA construct design. In Benchling, I assembled and annotated a complete cotton expression construct containing the SCFP fiber-specific promoter, start codon, optimized DFR coding sequence, 5’ UTR, 6×His tag, stop codon and NOS terminator. Finally, I utilized the process of designing a Twist order to validate whether the construct was technically synthesizable. After exporting the annotated plasmid sequence from Benchling, I uploaded it to Twist Bioscience and selected the pTwist Amp High Copy vector. The successful order validation confirmed that the construct met synthesis requirements and could theoretically be manufactured for laboratory testing.
Challenges
One unexpected challenge during the validation process was realizing that increasing DFR expression alone may not automatically result in visibly red, purple or blue cotton fibers. Although Gossypium hirsutum naturally contains the DFR gene, cotton fibers do not normally have any anthocyanin pigmentation, which suggests that additional regulatory factors or pathway limitations are involved. This made it difficult to predict whether codon optimization by itself would be sufficient to significantly enhance pigment production. To address this limitation, I compared DFR sequences from multiple highly pigmented plant species and considered the possibility of testing alternative DFR variants from plants such as grapes or strawberries in future experiments. A further limitation was that the project remained computational and theoretical due to the lack of access to a wet laboratory. Although the construct was successfully validated through Twist Bioscience and assembled digitally in Benchling, I could not experimentally test transformation efficiency, pigment intensity, or unintended physiological effects in real cotton plants.
Analysis
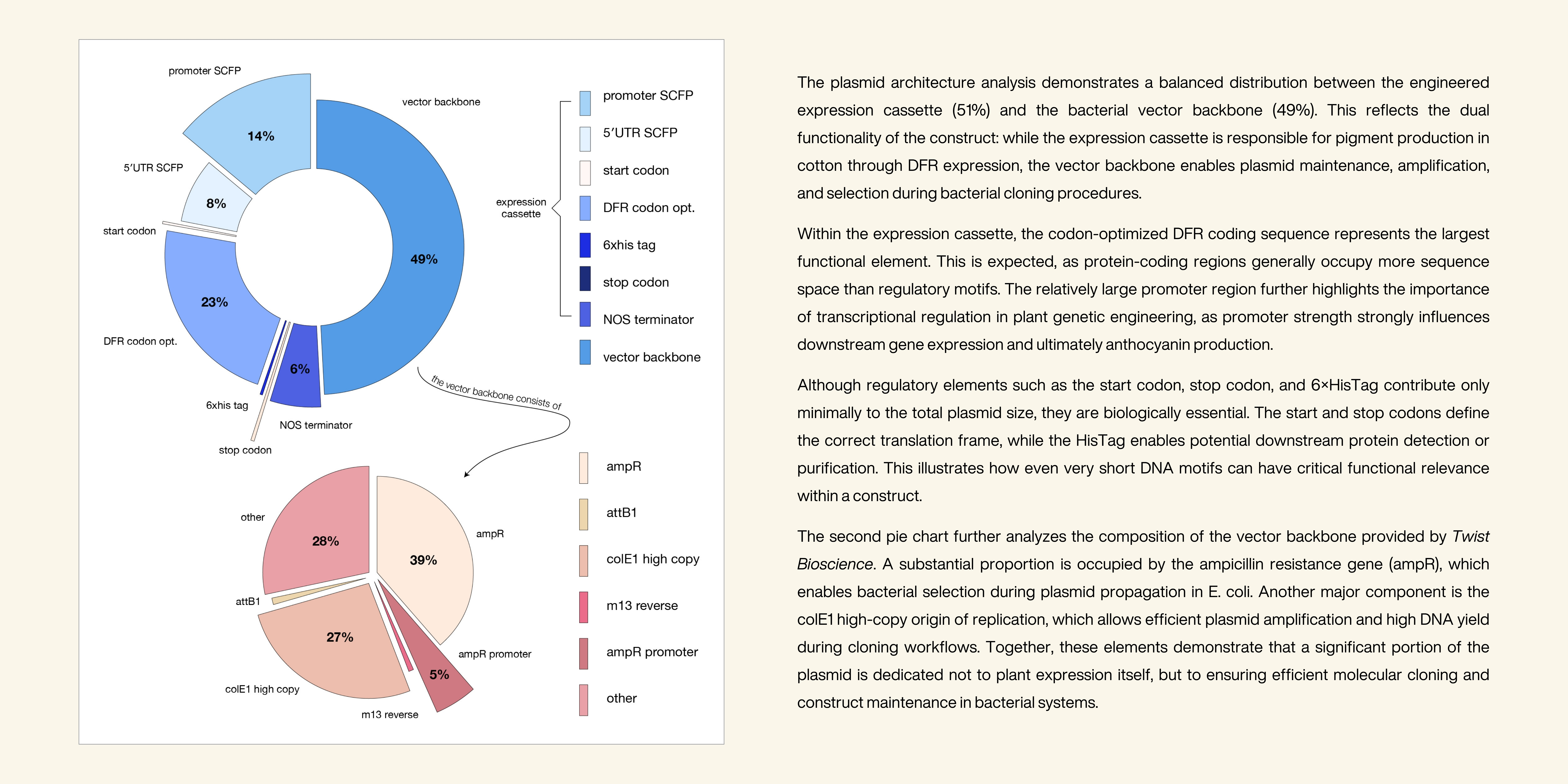
Conclusion
This project explored the possibility of rethinking textile coloration by shifting pigment production from an external industrial process to an intrinsic biological property of the material itself. By focusing on the anthocyanin biosynthesis pathway in Gossypium hirsutum, the project demonstrated that cotton already possesses the biological machinery required for pigment production, but that this capability is currently restricted to stress responses rather than fiber development. Through literature research, sequence analysis, codon optimization and plasmid design, a theoretically functional genetic construct for DFR expression in cotton was developed and digitally validated. The successful validation of the construct through Twist Bioscience confirmed that the designed sequence is technically synthesizable and compatible with standard molecular cloning systems. The project further highlighted the importance of regulatory design in plant synthetic biology. The use of a fiber-specific promoter, codon optimization strategies and established regulatory elements illustrated how gene expression can be computationally optimized to potentially improve anthocyanin production in cotton fibers. At the same time, the work demonstrated that even naturally occurring genes can be redesigned to achieve higher predicted expression efficiency, emphasizing the role of codon usage and regulatory architecture in synthetic construct design. Beyond the technical aspects, the project positioned genetically engineered pigmentation within a broader environmental and ethical context. Rather than treating wastewater after pollution has occurred, the concept proposes a preventative strategy that could reduce reliance on chemical dyes, water-intensive finishing processes and toxic wastewater generation. The use of hydroponic greenhouse systems additionally addressed biosafety concerns by providing a controlled cultivation environment that minimizes ecological exposure and supports responsible experimentation with genetically modified plants. Although the project remains conceptual and no biological transformation experiments were conducted, it demonstrates the possibility of intrinsically colored cotton fibers as a more sustainable alternative to conventional textile dyeing. Hopefully, one day it can be tested more thorougly within a lab environment.
Supply List and Budget
Since I am a non in-person comitted listener, my project is only conceptual and no budget is needed. My Twist Bioscience plasmid order would be $288.22.
References
Dhakal, K., Julkowska, M. M., Shekoofa, A. (2026) A hydroponic approach to assess the morpho-physiological responses of cotton cultivars under varying vapor pressure deficit conditions. Frontiers in Plant Science, 17:1751642 ↩︎
Natalio, F., Tahir M. N., Friedrich, N. et al. (2016) Structural analysis of Gossypium hirsutum fibers grown under greenhouse and hydroponic conditions. Journal of Structural Biology, http://dx.doi.org/10.1016/j.jsb.2016.03.005 ↩︎
Tan, J., Tu, L., Deng, F. et al. (2013) A Genetic and Metabolic Analysis Revealed that Cotton Fiber Cell Development Was Retarded by Flavonoid Naringenin, Plant Physiology, 162, 86-95 ↩︎
Chandler, S., Tanaka, Y. (2007) Genetic Modification in Floriculture. Critical Reviews in Plant Sciences, 26:4, 169-197, https://doi.org/10.1080/07352680701429381 ↩︎ ↩︎
Katsumoto, Y., Fukuchi-Mizutani, M., Fukui, Y. et al. (2007) Engineering of the Rose Flavonoid Biosynthetic Pathway Successfully Generated Blue-Hued Flowers Accumulating Delphinidin, Plant and Cell Physiology, 48:11, 1589–1600. https://doi.org/10.1093/pcp/pcm131 ↩︎
Parmar, M. S., Sharma, R. P. (2002) Development of various colours and shades in naturally coloured cotton fabrics. Indian Journal of Fibre & Textile Research, 27, 397-407 ↩︎
Gě, Q., Cūi, Y., Lǐ, J. et al. (2020) Disequilibrium evolution of the Fructose-1,6-bisphosphatase gene family leads to their functional biodiversity in Gossypium species. BMC Genomics 21, 379. https://doi.org/10.1186/s12864-020-6773-z ↩︎
Shi, S., Tang, R., Hao, X. et al. (2024) Integrative Transcriptomic and Metabolic Analyses Reveal That Flavonoid Biosynthesis Is the Key Pathway Regulating Pigment Deposition in Naturally Brown Cotton Fibers. Plants, 13, 2028. https://doi.org/10.3390/plants13152028 ↩︎ ↩︎
Pinnika, G., Thyssen, G. N., Madison, C. A. et al. (2025) The brown fiber phenotype in cotton line SA-40 is linked to a missing Ty3-like retrotransposon upstream of the GhTT2_A07. Frontiers in Plant Science, 16:1668965. doi: 10.3389/fpls.2025.1668965 ↩︎
Periyasamy, A. P. (2025) Textile Dyes in Wastewater and its Impact on Human and Environment: Focus on Bioremediation, Water Air Soil Pollut, 236:562 https://doi.org/10.1007/s11270-025-08204-7 ↩︎
Xie, D., Jackson, L. A., Cooper, J. D. (2004) Molecular and Biochemical Analysis of Two cDNA Clones Encoding Dihydroflavonol-4-Reductase from Medicago truncatula. Plant Physiology, 134, 979–994 ↩︎
Lewis, J. A., Zhang, B., Harza, R. et al. (2023) Structural Similarities and Overlapping Activities among Dihydroflavonol 4-Reductase, Flavanone 4-Reductase, and Anthocyanidin Reductase Offer Metabolic Flexibility in the Flavonoid Pathway. International Journal of Molecular Sciences, 24, 13901. https://doi.org/10.3390/ijms241813901 ↩︎
National Library of Medicine (2025). National center for biotechnology information. Nih.gov. Available at: https://www.ncbi.nlm.nih.gov/. ↩︎ ↩︎
Atella, A. L., Grossi-de-Sá, M. F., Alves-Ferreira, M. (2023) Cotton promoters for controlled gene expression. Electronic Journal of Biotechnology, 26, 1-12, https://doi.org/10.1016/j.ejbt.2022.12.002 ↩︎
Liliana, V. U.,Tatiana, C. N., Ricardo, P. C. et al. (2023) Plant Promoters: Their Identification, Characterization, and Role in Gene Regulation. Genes, 14, 1226. https://doi.org/10.3390/genes14061226 ↩︎
Yaqoob, A., Ali Shahid, A. , Salisu, I. B. et al. (2020) Comparative analysis of Constitutive and fiber-specific promoters under the expression pattern of Expansin gene in transgenic Cotton. PLOS ONE, 15(3):e0230519. https://doi.org/10.1371/journal.pone.0230519 ↩︎
Mayr, C. (2019) What Are 3′ UTRs Doing? Cold Spring Harbor Perspectives in Biology, 11:a034728 doi: 10.1101/cshperspect.a034728 ↩︎
Kimple, M. E., Brill, A. L., Pasker, R.L. (2013) Overview of affinity tags for protein purification. Curr Protoc Protein Sci, ;73:9.9.1-9.9.23. doi: 10.1002/0471140864.ps0909s73 ↩︎
Angenon, G., Van Montagu, M., Depicker, A. (1990). Analysis of the stop codon context in plant nuclear genes. , 271(1-2), 144–146. doi:10.1016/0014-5793(90)80392-v ↩︎