Phase 3: CTP Junction Design & SPP Cleavage Verification
Subcellular Targeting and Chloroplast Transit Peptide Engineering:
1. Selection of Chloroplast Transit Peptides
To improve targeting efficiency and avoid using repeated sequences, three different plant CTPs were selected:
- RbcS CTP: is derived from the Rubisco small subunit, one of the most abundant proteins in the chloroplast, and is widely used as a strong and reliable targeting signal.
- Fer2 CTP: comes from Ferredoxin-2, a chloroplast protein involved in electron transfer during photosynthesis, and is known for efficient import into the chloroplast stroma.
- RecA CTP: is derived from a chloroplast-localized RecA protein, which plays a role in DNA repair and maintenance within the chloroplast, and provides an alternative targeting signal with a different sequence composition.
These CTPs are derived from naturally chloroplast-targeted plant proteins (Arabidopsis thaliana) and are known to efficiently direct proteins into the chloroplast. Instead of using the same CTP for all seven genes, different peptides were intentionally distributed across the CODH subunits.
2. Fusion Design and Junction Engineering
Each CODH protein was fused to a CTP at its N-terminus. To make sure the protein folds correctly after cleavage, the fusion included the first 60 amino acids of each CODH protein, which were obtained using the ExPASy ProtParam tool.
A very important step was designing the junction between the CTP and the CODH protein. This region was carefully modified to include a cleavage motif recognized by the chloroplast enzyme responsible for removing the transit peptide:
(Val/Ile)-X-(Ala/Cys) ↓ Ala
To create this motif, small changes were made at the end of the CTP sequence as showed in the following sequences:
- RbcS CTP Sequence: MASSMLSSATMVASPAQATMVAPFNGLKSSAAFPATRKANNDITSITSNGGRVN(+AA)
- Fer2 CTP Sequence: MASTALSSAIVGTSFIRRSPAPISLRSLPSANTQSLFGLKSGTARGGRVTA(M–>A)
- RecA CTP Sequence: MDSQLVLSLKLNPSFTPLSPLFPFTPCSSFSPSLRFSSCYSRRLYSPVTVYA(+A)
This allowed a smooth transition between the CTP and the CODH protein while keeping both targeting and protein structure intact.
3. In Silico Validation of Targeting and Cleavage
All fusion sequences were analyzed using TargetP 2.0 to check two things:
- Whether the proteins are correctly targeted to the chloroplast
- Where the CTP is predicted to be cleaved
![]()
![]()
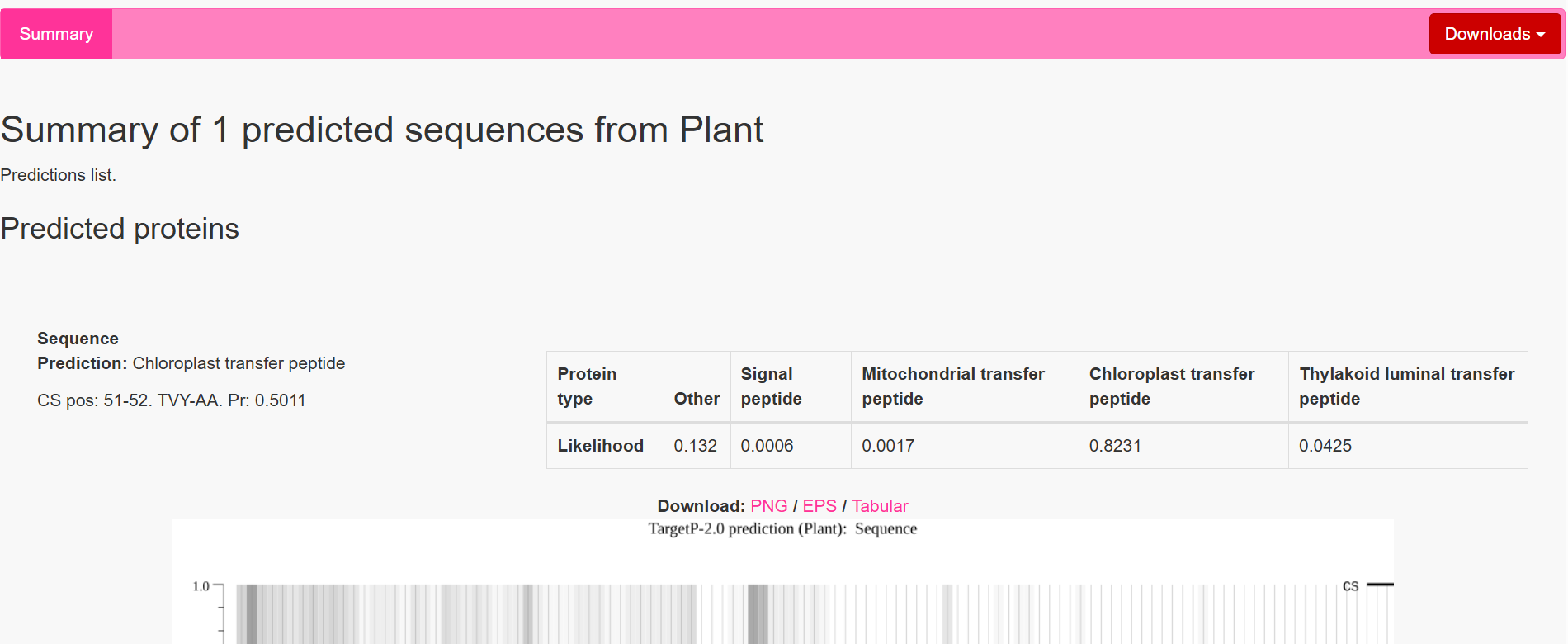
The results showed that all seven proteins are predicted to be targeted to the chloroplast, which confirms that the CTPs are working correctly:
.png)
.png)
.png)
.png)
.png)
.png)
.png)
Summary of the results:
| Gene | CTP Source | Cleavage Site (CS Position) | Junction Motif (CTP → CODH) | Cleavage Probability (Pr) | Prediction |
|---|---|---|---|---|---|
| coxD | RbcS | 55–56 | VNA ↓ AM | 0.5216 | Chloroplast transfer peptide |
| coxE | Fer2 | 51–52 | VTA ↓ AM | 0.3172 | Chloroplast transfer peptide |
| coxF | RecA | 51–52 | TVY ↓ AA | 0.4989 | Chloroplast transfer peptide |
| coxG | RbcS | 55–56 | VNA ↓ AM | 0.5923 | Chloroplast transfer peptide |
| coxL | RbcS | 55–56 | VNA ↓ AM | 0.4842 | Chloroplast transfer peptide |
| coxM | Fer2 | 51–52 | VTA ↓ AM | 0.7188 | Chloroplast transfer peptide |
| coxS | RecA | 51–52 | TVY ↓ AA | 0.5011 | Chloroplast transfer peptide |
Interpretation of Results
Overall, the results indicate successful design of functional targeting signals for all CODH subunits:
All constructs were confidently predicted as chloroplast-targeted proteins, confirming that the added CTPs are functional. The cleavage sites align well with the engineered junction motifs, demonstrating that the proteins are likely to be correctly processed after import.
The coxM fusion showed the highest cleavage probability (Pr = 0.7188), indicating highly efficient targeting and processing. Other subunits showed moderate probabilities (around 0.48–0.59), which are still within acceptable ranges for functional targeting. The coxE fusion presented a lower probability (Pr = 0.3172). Although this suggests potentially less efficient cleavage, the sequence still satisfies the required motif and is expected to remain functional, as variability in cleavage efficiency is common in heterologous systems.
Most constructs showed cleavage occurring exactly at the designed motif, typically between amino acid positions 51–56, depending on the transit peptide used.
However, a notable observation was made for two constructs, coxF and coxS, where the predicted cleavage site occurred slightly upstream of the engineered junction, specifically just before the designed alanine-alanine region rather than directly within it.
This slight variation in cleavage position is consistent with the known behavior of the chloroplast Stromal Processing Peptidase. Rather than recognizing a single fixed sequence, the enzyme identifies a broader structural and sequence context, which allows for some flexibility in the exact cleavage position. As a result, small shifts of one or two amino acids relative to the designed motif are commonly observed in both native and engineered proteins.
In this case, although the cleavage in coxF and coxS occurs marginally earlier than expected, it remains within a functionally acceptable region. The resulting mature proteins retain nearly identical N-terminal sequences and are not expected to lose any essential structural or functional elements. Importantly, the targeting prediction remains strong, confirming that the proteins are still efficiently directed to the chloroplast.
Therefore, this variability does not compromise the overall design. All fusion constructs are considered valid, and no redesign was required. Instead, this observation reflects the inherent flexibility of chloroplast protein processing and further validates the robustness of the engineered system.
Objective
Subcellular targeting is a critical step in synthetic biology when expressing proteins in a new host organism. In plant cells, proteins must be directed to the correct organelle in order to function properly. This is especially important for metabolic pathways that depend on specific cellular environments.
In this project, the seven proteins forming the Carbon Monoxide Dehydrogenase (CODH) system originate from a bacterium. However, in plant cells, these proteins need to function inside the chloroplast, where photosynthesis occurs and where the produced CO₂ can be directly reused.
Bacterial proteins do not naturally contain signals that allow them to enter plant organelles. As a result, if they are expressed without modification, they will remain in the cytosol, where they may not fold correctly, may not interact properly with other subunits, and may fail to form a functional enzyme complex.
To solve this problem, each CODH protein must be fused to a chloroplast transit peptide (CTP). These short sequences are naturally found in plant proteins and act as targeting signals that guide newly synthesized proteins into the chloroplast. Once the protein reaches the chloroplast, the transit peptide is cleaved, releasing the mature protein in its functional form.
Sources:
- An optimized transit peptide for effective targeting of diverse foreign proteins into chloroplasts in rice | Scientific Reports. (n.d.). Retrieved May 5, 2026, from https://www.nature.com/articles/srep46231
- Caspari, O. D. (2022). Transit Peptides Often Require Downstream Unstructured Sequence for Efficient Chloroplast Import in Chlamydomonas reinhardtii. Frontiers in Plant Science, 13. https://doi.org/10.3389/fpls.2022.825797
- Caspari, O. D., Garrido, C., Law, C. O., Choquet, Y., Wollman, F.-A., & Lafontaine, I. (2023). Converting antimicrobial into targeting peptides reveals key features governing protein import into mitochondria and chloroplasts. Plant Communications, 4(4), 100555. https://doi.org/10.1016/j.xplc.2023.100555
- Chung, B. K.-S., & Lee, D.-Y. (2012). Computational codon optimization of synthetic gene for protein expression. BMC Systems Biology, 6, 134. https://doi.org/10.1186/1752-0509-6-134
- Codon Adaptation Index. (2024). In Wikipedia. https://en.wikipedia.org/w/index.php?title=Codon_Adaptation_Index&oldid=1254549471
- Dietel, A.-K., Merker, H., Kaltenpoth, M., & Kost, C. (2019). Selective advantages favour high genomic AT-contents in intracellular elements. PLoS Genetics, 15(4), e1007778. https://doi.org/10.1371/journal.pgen.1007778
- Lee, S., Weon, S., Lee, S., & Kang, C. (2010). Relative Codon Adaptation Index, a Sensitive Measure of Codon Usage Bias. Evolutionary Bioinformatics Online, 6, 47–55. https://doi.org/10.4137/ebo.s4608
- Li, Q., Luo, Y., Sha, A., Xiao, W., Xiong, Z., Chen, X., He, J., Peng, L., & Zou, L. (2023). Analysis of synonymous codon usage patterns in mitochondrial genomes of nine Amanita species. Frontiers in Microbiology, 14. https://doi.org/10.3389/fmicb.2023.1134228
- Monjezi, Z., Rooshanfekr, H. allah, Nazari, M., Salabi, F., & Tabandeh, M. R. (2024). Codon optimization of voraxin α sequence enhances the immunogenicity of a recombinant vaccine against Hyalomma anatolicum infestation in rabbits. Veterinary Immunology and Immunopathology, 275, 110817. https://doi.org/10.1016/j.vetimm.2024.110817
- Puigbò, P., Bravo, I. G., & Garcia-Vallve, S. (2008). CAIcal: A combined set of tools to assess codon usage adaptation. Biology Direct, 3, 38. https://doi.org/10.1186/1745-6150-3-38
- Richter, S., & Lamppa, G. K. (1999). Stromal Processing Peptidase Binds Transit Peptides and Initiates Their Atp-Dependent Turnover in Chloroplasts. The Journal of Cell Biology, 147(1), 33–44. https://doi.org/10.1083/jcb.147.1.33
- Supek, F., & Šmuc, T. (2010). On Relevance of Codon Usage to Expression of Synthetic and Natural Genes in Escherichia coli. Genetics, 185(3), 1129–1134. https://doi.org/10.1534/genetics.110.115477
- Thagun, C., Odahara, M., Kodama, Y., & Numata, K. (2024). Identification of a highly efficient chloroplast-targeting peptide for plastid engineering. PLOS Biology, 22(9), e3002785. https://doi.org/10.1371/journal.pbio.3002785
- Willems, T., Hectors, W., Rombaut, J., De Rop, A.-S., Goegebeur, S., Delmulle, T., De Mol, M. L., De Maeseneire, S. L., & Soetaert, W. K. (2023). An exploratory in silico comparison of open-source codon harmonization tools. Microbial Cell Factories, 22, 227. https://doi.org/10.1186/s12934-023-02230-y