Week 11 HW: Bioproduction & Cloud Labs
Part A: The 1,536 Pixel Artwork Canvas | Collective Artwork
1. My contribution
Unfortunately, I was not able to contribute a pixel to the collective artwork, as I was in the middle of midterm exams at my university during that period, which limited my availability to participate.
2. What I liked about the project
I really liked the project because of its biological foundation and particularly its connection to cell-free fluorescent protein optimization and how it was used for a global pixel artwork designed by HTGAA students.
3. What could be improved for next year
For future versions, it could be interesting to include a live chat feature so participants can coordinate in real time and create more elaborate and intentional designs. Additionally, increasing the number of pixels beyond the 1,536 used in this edition could allow for more detailed and realistic compositions.
Update :) !:

There was a second part of the Pixel Artwork where I was able to contribute to the design of the bacterium shown on the right. Later, when I checked the design again, I noticed that other classmates had contributed to improving it as well, such as adding a smile and giving it an outline :D

(Also, haappy to be one of the main contributors this time :] )
Figure: Building genomes workflow and design overview.
Part B: Cell-Free Protein Synthesis | Cell-Free Reagents
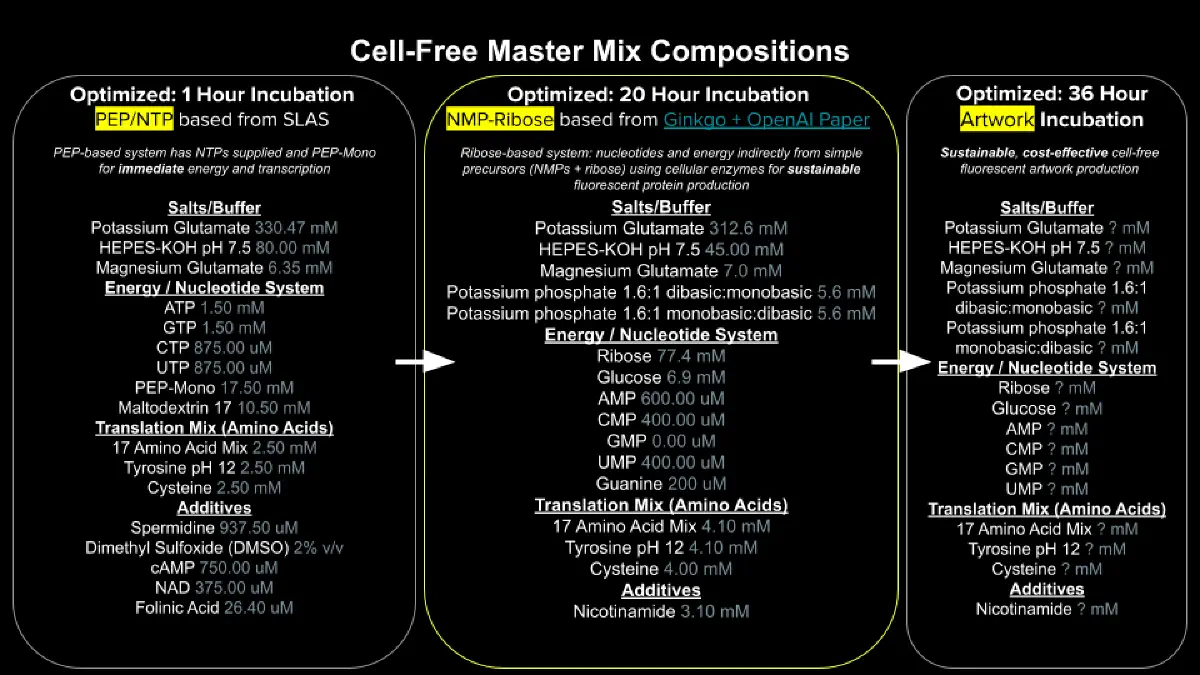
- Referencing the cell-free protein synthesis reaction composition (the middle box outlined in yellow on the image above, also listed below), provide a 1-2 sentence description of what each component’s role is in the cell-free reaction.
E. coli Lysate
BL21 (DE3) Star Lysate (includes T7 RNA Polymerase)
The lysate provides the essential molecular machinery required for transcription and translation, including ribosomes, metabolic enzymes, cofactors, and tRNAs. The presence of T7 RNA polymerase enables efficient transcription of target genes under T7 promoter control.
Salts / Buffer
- Potassium Glutamate
Maintains ionic strength and mimics intracellular conditions, thereby stabilizing macromolecular interactions and supporting enzymatic activity.
- HEPES-KOH pH 7.5¨
Serves as a buffering agent to maintain a stable pH, which is critical for optimal enzyme function during transcription and translation.
- Magnesium Glutamate
Functions as an essential cofactor for ribosomes and polymerases, playing a key role in both transcriptional and translational processes.
- Potassium Phosphate Monobasic / Dibasic
Contributes to buffering capacity and provides phosphate ions necessary for nucleotide metabolism and energy transfer reactions.
Energy / Nucleotide System
- Ribose
Acts as a precursor for nucleotide biosynthesis, supporting sustained RNA production over extended reaction times.
- Glucose
Serves as a metabolic energy source, enabling ATP regeneration through endogenous enzymatic pathways present in the lysate.
- AMP, CMP, GMP, UMP
Provide nucleotide monophosphates that can be phosphorylated into their corresponding triphosphates, which are required substrates for RNA synthesis.
- Guanine
Functions as a precursor in nucleotide salvage pathways, allowing for the biosynthesis of GMP and subsequently GTP for transcription.
Translation Mix (Amino Acids)
- 17 Amino Acid Mix
Supplies the majority of amino acids required for protein synthesis during translation.
- Tyrosine
Provided separately due to solubility and stability constraints; essential for incorporation into nascent polypeptides.
- Cysteine
Added separately due to its susceptibility to oxidation; plays a critical role in protein structure through disulfide bond formation.
Additives
- Nicotinamide
Supports redox balance and enzymatic activity by contributing to NAD⁺-dependent metabolic processes within the reaction. Backfill
- Nuclease-Free Water
Used to adjust the final reaction volume while preventing nucleic acid degradation.
- Describe the main differences between the 1-hour optimized PEP-NTP master mix and the 20-hour NMP-Ribose-Glucose master mix shown in the Google Slide above. (2-3 sentences)
The 1-hour PEP-NTP system relies on the direct addition of high-energy phosphate donors and nucleotide triphosphates, enabling rapid and high-yield protein synthesis within a short time frame. However, this approach is limited by the rapid depletion of energy substrates and accumulation of inhibitory byproducts.
In contrast, the 20-hour NMP-ribose-glucose system employs a metabolically sustained strategy in which substrates such as ribose and glucose support continuous nucleotide regeneration and ATP production. This results in prolonged reaction stability and sustained protein expression over extended periods.
- Bonus question: How can transcription occur if GMP is not included but Guanine is?
Transcription can still occur in the absence of externally supplied GMP because guanine can be converted into GMP through endogenous nucleotide salvage pathways present in the lysate. The resulting GMP can then be phosphorylated to GTP, which serves as the direct substrate for RNA polymerase during transcription.
References:
Carlson, E. D., Gan, R., Hodgman, C. E., & Jewett, M. C. (2012). Cell-free protein synthesis: Applications come of age. Biotechnology Advances, 30(5), 1185–1194. https://doi.org/10.1016/j.biotechadv.2011.09.016
Swartz, J. R. (2012). Transforming biochemical engineering with cell-free biology. AIChE Journal, 58(1), 5–13. https://doi.org/10.1002/aic.13701
Part C: Planning the Global Experiment | Cell-Free Master Mix Design
1. Given the 6 fluorescent proteins we used for our collaborative painting, identify and explain at least one biophysical or functional property of each protein that affects expression or readout in cell-free systems. (Hint: options include maturation time, acid sensitivity, folding, oxygen dependence, etc) (1-2 sentences each)
1. sfGFP: Rapid Folding
sfGFP (superfolder GFP) can fold and form a functional chromophore in under 10 minutes, making it one of the fastest-maturing fluorescent proteins available. This is largely due to key stabilizing mutations like S30R and Y39N, which reinforce the β-barrel structure and improve global stability, meaning sfGFP begins producing fluorescence signal almost immediately after translation in a cell-free system, ideal for short incubations.
2. mRFP1: Incomplete Maturation / Low Quantum Yield
mRFP1 is reported to be a somewhat slowly-maturing monomer with low acid sensitivity; although it matures more than 10 times faster than its precursor DsRed, it still has a lower extinction coefficient, quantum yield, and photostability. In a cell-free context, this means a meaningful fraction of translated mRFP1 may remain non-fluorescent at any given time point, limiting the overall signal readout compared to what the protein concentration would predict.
3. mKO2: High Oxygen Dependence
mKO2 has a comparatively high dependence on oxygen tension for chromophore maturation, the oxygen tension at which 50% of fluorescence-positive cells is lost (pO₂·50) is ~0.9% for mKO2, and its kinetics of fluorescence recovery after reoxygenation are much slower than for greener variants like mAG. In a sealed or oxygen-limited cell-free reaction, this strong oxygen requirement means mKO2 may substantially underperform, as the chromophore oxidation step is rate-limiting and difficult to rescue once oxygen is depleted.
4. mTurquoise2: Exceptionally High Quantum Yield
mTurquoise2 has the highest quantum yield measured for a monomeric fluorescent protein, along with fast maturation, high photostability and a long mono-exponential fluorescence lifetime. It is also reported to be a rapidly-maturing monomer with very low acid sensitivity, making it one of the most reliable reporters in cell-free systems, its high quantum yield directly translates into more photons emitted per molecule, maximizing signal even at moderate expression levels.
5. mScarlet-I: Accelerated Maturation at the Cost of Quantum Yield
The single amino acid substitution T74I in mScarlet-I results in a marked maturation acceleration in cells, but at the cost of a moderate decrease in fluorescence quantum yield (0.54) and fluorescence lifetime (3.1 ns), though both values are still higher than those of all previously engineered bright mRFPs. This trade-off is particularly relevant in cell-free incubations: faster chromophore maturation is advantageous because it means fluorescence appears sooner, but the reduced quantum yield means the peak brightness will be lower than the parental mScarlet.
6. Electra2: Context-Dependent Brightness and Aggregation Risk
Electra2 is a monomeric blue fluorescent protein developed through hierarchical screening in bacterial and mammalian cells, optimized for intracellular brightness in a spectral range underserved by previous BFPs. However, its brightness performance varies significantly across expression systems — Electra2 outperforms mTagBFP2 in zebrafish but is dimmer in mouse brain under two-photon excitation, and there is a higher tendency for Electra1 (its close relative) to form puncta in neurons in vivo. In a cell-free system lacking the chaperone and quality-control machinery of living cells, Electra2’s tendency toward aggregation at higher concentrations could reduce functional fluorescent protein yield.
2. Create a hypothesis for how adjusting one or more reagents in the cell-free mastermix could improve a specific biophysical or functional property you identified above, in order to maximize fluorescence over a 36-hour incubation. Clearly state the protein, the reagent(s), and the expected effect.
Protein: mKO2
Property to improve: Oxygen-dependent chromophore maturation
Hypothesis: Because mKO2 exhibits the strongest dependence on oxygen tension of all six proteins used, its fluorescence readout in a 36-hour cell-free incubation is likely limited by progressive oxygen depletion in the sealed reaction volume. As demonstrated in PURE system studies, oxygen is not only consumed during chromophore maturation of fluorescent proteins, but also by ATP regeneration pathways. For example, the pyruvate oxidase-based ATP regeneration system requires molecular oxygen to generate acetyl phosphate from pyruvate, creating direct competition for dissolved O₂.
Proposed adjustment: Supplement the mastermix with catalase (which regenerates O₂ from H₂O₂ produced during oxidative reactions) and use open or semi-permeable reaction vessels rather than sealed tubes to allow passive oxygen diffusion throughout the 36-hour incubation. This strategy is supported by studies in cell-free fluorescence fluctuation spectroscopy, where a droplet format was specifically designed to ensure sufficient oxygenation for chromophore maturation. The expected effect is a sustained supply of O₂ across the incubation window, enabling mKO2’s slower oxidation kinetics to run to completion and significantly increasing total fluorescence yield compared to an oxygen-depleted closed system.
Sources:
Pedelacq, J.-D., Cabantous, S., Tran, T., Terwilliger, T. C., & Waldo, G. S. (2006). Engineering and characterization of a superfolder green fluorescent protein. Nature Biotechnology, 24(1), 79–88. https://www.nature.com/articles/nbt1172
Campbell, R. E., Tour, O., Palmer, A. E., Steinbach, P. A., Baird, G. S., Zacharias, D. A., & Tsien, R. Y. (2002). A monomeric red fluorescent protein. Proceedings of the National Academy of Sciences, 99(12), 7877–7882. https://www.pnas.org/doi/10.1073/pnas.082243699
Kagawa, W., Aida, T., Oguro, T., & Iida, R. (2012). Differential dependence on oxygen tension during the maturation process between monomeric Kusabira Orange 2 and monomeric Azami Green expressed in HeLa cells. Biochemical and Biophysical Research Communications, 422(2), 267–272. https://www.sciencedirect.com/science/article/abs/pii/S0006291X1200784X
Goedhart, J., von Stetten, D., Noirclerc-Savoye, M., Lelimousin, M., Joosen, L., Hink, M. A., van Weeren, L., Gadella, T. W. J., & Royant, A. (2012). Structure-guided evolution of cyan fluorescent proteins towards a quantum yield of 93%. Nature Methods, 9(3), 259–261. https://pubmed.ncbi.nlm.nih.gov/22434194/
Bindels, D. S., Haarbosch, L., van Weeren, L., Postma, M., Wiese, K. E., Mastop, M., Aumonier, S., Gotthard, G., Royant, A., Hink, M. A., & Gadella, T. W. J. (2017). mScarlet: a bright monomeric red fluorescent protein for cellular imaging. Nature Methods, 14(1), 53–56. https://www.nature.com/articles/nmeth.4074
Papadaki, S., Bhatt, D., & Bhatt, P. (2022). Dual-expression system for blue fluorescent protein optimization. Scientific Reports, 12, 10081. https://www.nature.com/articles/s41598-022-13214-0
Dunsing, V., Petrov, E. P., & Schwille, P. (2012). Chromophore maturation and fluorescence fluctuation spectroscopy of fluorescent proteins in a cell-free expression system. Biophysical Journal, 102(11), 2536–2545. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3367886/
Lavickova, B., & Maerkl, S. J. (2025). ATP regeneration from pyruvate in the PURE system. ACS Synthetic Biology. https://pubs.acs.org/doi/10.1021/acssynbio.4c00697
3. The second phase of this lab will be to define the precise reagent concentrations for your cell-free experiment. You will be assigned artwork wells with specific fluorescent proteins and receive an email with instructions this week (by April 24)
Assigned protein: mKO2 (self-assigned, no email received)
Rationale
Based on the biophysical analysis in question 1 and the hypothesis in question 2, mKO2 was selected as the target protein for reagent optimization. Its primary limitation in a 36-hour cell-free incubation is its strong dependence on molecular oxygen for chromophore maturation, a resource that becomes progressively limiting in a closed 20 μL reaction as both the ATP regeneration machinery and the chromophore oxidation step compete for dissolved O₂.
The custom 2 μL supplement makes two small but targeted adjustments over the default preset, with the remaining volume completed with nuclease-free water:
1. Potassium Phosphate Dibasic (+25 nL): A modest increase above the preset 5.625 mM. Phosphate is a direct substrate in the oxidative ATP regeneration pathway; maintaining slightly higher phosphate availability over a 36-hour incubation helps sustain energy production and reduces metabolic competition for oxygen, leaving more dissolved O₂ available for mKO2 chromophore maturation.
2. Magnesium Glutamate (+25 nL): A small increase above the preset 6.975 mM. Mg²⁺ is an essential cofactor for RNA polymerase and ribosomes, and a modest bump is expected to improve translational yield, producing a larger pool of mKO2 polypeptide that can mature fluorescently as oxygen becomes available. The increase was kept minimal to avoid exceeding ~10 mM, above which excess Mg²⁺ can inhibit cell-free transcription-translation.
3. Nuclease-Free Water (1950 nL): Fills the remainder of the 2 μL supplement volume, keeping the total reaction at 20 μL without introducing unintended osmotic or ionic effects.
Expected Outcome: These conservative adjustments aim to sustain energy regeneration and translational output over the full 36-hour window, maximizing the total pool of matured mKO2 fluorophore. The hypothesis is that even small improvements in phosphate buffering and Mg²⁺ availability will shift the limiting factor away from energy depletion, allowing the reaction to run closer to its theoretical maximum fluorescence output given mKO2’s inherently slower oxidation kinetics.
Reagent Supplement JSON
(Total supplement volume: 2000 nL = 2 μL, within the assigned limit)
Additional Well: Electra2 - “Slow Burn” Strategy
Target Reporter:
Electra2
Operational Design:
Delayed Maturation / Late-Peak Pixel
Rationale
Electra2 was selected as a second experimental well to explore a dimension not emphasized in other designs within the collaborative painting: fluorescence persistence and delayed maturation dynamics rather than rapid early brightness.
Unlike fast-folding Aequorea-derived proteins such as sfGFP or mTurquoise2, Electra2 originates from a coral fluorescent protein lineage (eqFP611 via mRuby3-derived engineering), which undergoes comparatively slower and more complex chromophore maturation. This creates the possibility that fluorescence accumulation continues later into the 36-hour incubation window, even after other wells have plateaued.
To support this delayed maturation profile, the reaction environment is biased toward maximum pH stability using elevated HEPES buffering capacity. During long incubations, cell-free reactions gradually accumulate acidic metabolic byproducts that can reduce fluorescence intensity or prematurely quench partially matured chromophores. Increasing HEPES concentration helps preserve a near-optimal pH environment throughout the incubation period, extending the effective maturation window for Electra2.
Rather than maximizing instantaneous signal intensity, this design aims to produce a late-emerging fluorescence phenotype or a pixel that continues strengthening over time while faster systems stabilize or decay.
Reagents adjusted:
| Reagent | Preset | Set to |
|---|---|---|
| HEPES-KOH pH 7.5 | 45.000 mM | Increased |
| Nuclease-Free Water | balance | balance |
Expected Phenotype / Visual Outcome
A blue fluorescent pixel with delayed onset and prolonged signal persistence, serving as a temporal contrast element within the collaborative painting. The design demonstrates that fluorescence timing and maturation kinetics can be intentionally shaped in cell-free expression systems.
Reagent Supplement JSON
(Total supplement volume: 2000 nL = 2 μL)