Dia-form: Modifying the Biosilica Deposit Structure of Diatoms
Abstract
Clear glass has been crucial to the development of modern architecture, with windows and clear glazing being a major catalyst for indoor living. However this dependence on clear glass has also created a dependence on new material, from a specific and limited sand for glass making, as well as high energy use to fire and float form the sand into glass panes. This project aims at exploring a biological alternative to glass panes, as well as developing the scientific results that could point to future work that uses this biosilica for novel materials, both aggregated with construction waste and as a pure material out of diatom blooms. Diatoms are a type of algae that have silica cell walls called frustules, and these frustules form into intricate lacy, opalescant patterns as the colonies of algae grow. Cylindrotheca fusiformis is a marine diatom species that relies on proteins including silaffins for silicic acid polymerization. By modifying the proteins that are responsible for the diatom structure, this project opens up the mechanical properties of diatoms as a material, where structure is responsible for color expression and for potential material attachment and other characteristics for future projects. Initial work would be conducted in cell-free systems before moving into algal systems once again to work towards creating a ‘glass’ panel with shifting structural properties created through silaffin modifications.
Project Aims
Protein Modification Modify the Sil1p protein of Cylindrotheca fusiformis through cell-free systems. In this aim, I am interested in observing a change of the cell-free system after Sil1p modification from the original sequence. The modification would aim to either overexpress the polymerizing protein, or supress it and see the potential shifts that emerge from those DNA modifications. These shifts would be tested using florescent tagging and a colorimetric assay to check if the system worked, and whether there are notable changes between the modified proteins.
Algae Modification Modify a non-diatom, and non-frustule forming algae and hopefully, C.fusiformis itself, to have the Sil1p protein in place of their traditional cell wall proteins. This would be an exciting yet achievable way to observe the protein in an algae structure, focusing on addressing the challenge of modifying the diatoms without killing the cells.
(OLD AIM 2: Use CRISPR/Cas-9 or the gene gun (most likely CRISPR) to input the modified protein into the T.pseudonona THIS CHANGED AFTER FURTHER DIATOM MODIFICATION RESEARCH.)
Color Shift Modify the TpSil1/2 protein of C.fusiformis to refract light with a visibly different hue through structural modification. The overexpression or knockout of this gene can result in more or less silica deposition, resulting in an altered macropore structure, and thus modifying the light scattering by the physical structure of the frustules.
Rubble Attachment Further modifying the diatom structure to act as a bandaid between two pieces of glass, or two pieces of cement rubble. This takes the structural changes of the silica deposition to a new function, using it to support the use of hard-to-modify architectural waste for new architectural material. Can diatoms from silica patterns that attach onto surrounding objects/surfaces?
Section 3: Background
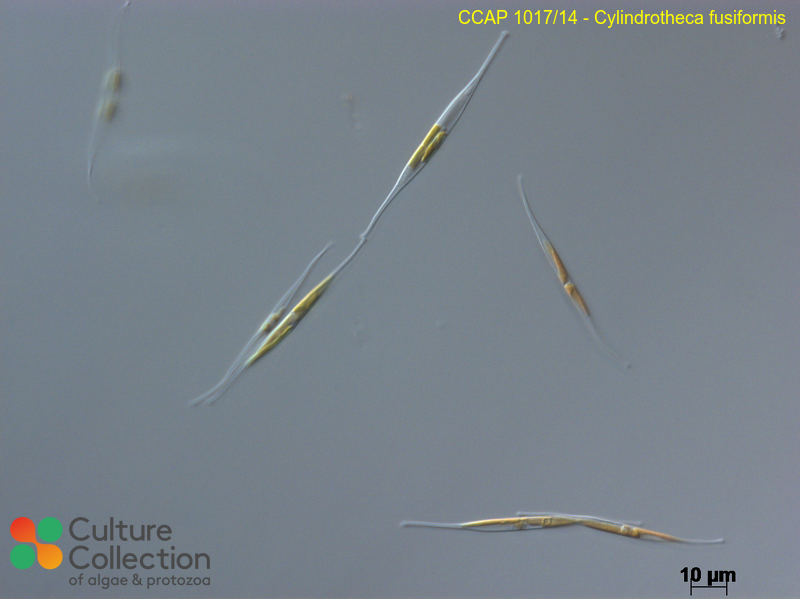
Diatoms are a unique class of algae that produce silica cell walls called frustules. These frustules have shown up in fossil records, indicating a continued presence of diatoms in our environment over millenia. They are also key organisms for carbon fixation, often looked into for biofuel production. However, by celebrating the unique silica structure of diatoms, this project brings a novel angle of bio-stained glass for architectural applictions to the ways that we may interact with diatoms. Diatoms have been used as aggregates to things like diatomaceous earth, pest control, and for biofuels for their lipid production. Glass production for glazing has created a heavy dependency on specific sand sources that are outsized in a world of skyscrapers. This project proposes an alternative to a sand-derived glass through diatoms. While this project does not suggest that every piece of glass should be made with diatoms from here on out, the production of even a single pane of glass, or single stained glass installation creates a unique condition of a species beyond its natural growning environment and of glass beyond its pure, inert, and uniform state.
Diatoms play an outsized role in oxygen production in the world, (Li et all 2025) producing around 20%, however its silica structure is more of the key focus for this project, with fossil records dating back to the Lower Cretascous Age (https://ucmp.berkeley.edu/chromista/diatoms/diatomfr.html) thanks to their hard silica walls. Kroger et al (1999) identified silaffin as a key protein in the polymerization of silicic acid from aquatic environments into the silica cell walls of diatoms. The R5 peptides were also identified for its repeted occurance within the protein, and its role as the nucleation catalyst for the silicic acid was also identified, acting like scaffolding for the silica to precipitate and harden (Sumper and Brunner 2008), and this peptide plays a key role in the pattern of the silica deposition. From these papers, I focused on the role of a single protein, Sil1p, and the R5 peptide to narrow the experiment to a key singular modification, while Tong et al (2021) speaks towards the importance of the environmental conditions of the silaffin for the growth and attachment, showing that the conditions for growth such as container surface may play a role in shaping the silica deposits beyond the protein modifitions. C.fusiformis is one of the few diatoms that are well studied at this time, and the modification of such a diatom could lead to future experiments into the potential for other modification-resistant organisms to modulate the built environment.
This bioglass approach offers a new way for glass to be considered in the built environment, and the macropore structure gives way for other organisms to occupy these panels in the urban environment. The inherent carbon fixation of the diatoms, and the continued environmental impact of the supported microorganisms like photosynthetic algae, or moss and lichen, can help mitigate the biodiversity depeltion of urban spaces that are often overbuilt in glass. They also provide a softened edge, breaking the flood-prone smooth surfaces with the pore structure. In a synthetic biology context, this project aims to modify a very rigid cell structure, contributing to the body of literature that looks to modifications for organisms that produce calcium and silica-rich structures that could be beneficial for synthetic bones, alternative fuels, and more. This project reframes the inertness of glass, and blurs the boundary between the permanent and temporary materials of the environment. If these panels could be created with living diatoms, their growth patterns would indicate environmental conditions like water stagnation and sun exposure, creating a panel with morphology that is hyper-local, something that architects have been attempting to immitate through computational simulations and programs for decades now, that a large research area of computational design is based off of.
Governance Concerns and Speculated Solutions
- Impact on local ecosystems: This design depends on the cultivation of a large quantity of diatoms, which can impact local water ecosystems if not contained in a proper manner.
To address this impact, this project speculates conditions similar to algal production for biofuel, however the unique silica deposits of diatoms make the container a more complex area for research. Aquarium-style tanks would be a good place to start, with wider tubes to account for silica deposits within the filtration system.
- Impact on other organisms: If these modified diatoms were to be introduced to an environment through the proposed architectural panels, is there a chance that the mutated diatoms can impact the other organisms in the ecosystems around it?
To address this potential issue, the diatoms will be studied at a petri dish scale tanks with model ecosystem samples, and the other organisms in these sample ecosystems will be tested compared to the control, unmodified diatom ecosystem to see whether there is an impact. Until those tests have a conclusion and beyond, the diatoms can be kiln fired to kill the diatoms in the panel before installation.
- Dependence on specific resources: This, like glass, is dependent on a narrow resource, creating issues for future scale up.
This would be addressed through lab-based cultivation and harvesting, creating an initial dependence on a wild organism before creating a contained synthetic growth system to harvest from.
Experimental Design, Techniques, Tools, and Technology
Aim 1’s focus of protein modification began with a study of the silaffin protein and the key peptides that shape it.
Benchling link to Aim 1 notes: https://benchling.com/s/etr-irtNof3IiIZ01CwszzmL?m=slm-b4xotTiSKGL4vUWfUjc9
Sil1p c.fusiformis sequence and protein structure without modification:
MKLTAIFPLLFTAVGYCAAQSIADLAAANLSTEDSKSAQLISADSSDDASDSSVESVDAASSDVSGSSVESVDVSGSSLESVDVSGSSLESVDDSSEDSEEEELRILSSKKSGSYYSYGTKKSGSYSGYSTKKSASRRILSSKKSGSYSGYSTKKSGSRRILSSKKSGSYSGSKGSKRRILSSKKSGSYSGSKGSKRRNLSSKKSGSYSGSKGSKRRILSSKKSGSYSGSKGSKRRNLSSKKSGSYSGSKGSKRRILSGGLRGSM
source: https://www.uniprot.org/uniprotkb?query=cylindrotheca+fusiformis
DNA reverse translation of the Sil1p protein sequence from https://www.bioinformatics.org/sms2/rev_trans.html
reverse translation of sp|Q9SE35|SIL1_CYLFU Silaffin-1 OS=Cylindrotheca fusiformis OX=2853 GN=SIL1 PE=1 SV=1 to a 795 base sequence of most likely codons.
atgaaactgaccgcgatttttccgctgctgtttaccgcggtgggctattgcgcggcgcagagcattgcggatctggcggcggcgaacctgagcaccgaagatagcaaaagcgcgcagctgattagcgcggatagcagcgatgatgcgagcgatagcagcgtggaaagcgtggatgcggcgagcagcgatgtgagcggcagcagcgtggaaagcgtggatgtgagcggcagcagcctggaaagcgtggatgtgagcggcagcagcctggaaagcgtggatgatagcagcgaagatagcgaagaagaagaactgcgcattctgagcagcaaaaaaagcggcagctattatagctatggcaccaaaaaaagcggcagctatagcggctatagcaccaaaaaaagcgcgagccgccgcattctgagcagcaaaaaaagcggcagctatagcggctatagcaccaaaaaaagcggcagccgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcaacctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcaacctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcggcggcctgcgcggcagcatg
codon optimized sequence for e.coli based cell-free systems (source: https://en.vectorbuilder.com/tool/codon-optimization/85f15f77-6cd8-42c7-a356-73014dc090f0.html)
ATGAAACTGACCGCAATTTTTCCGCTGCTGTTTACCGCCGTGGGCTATTGCGCCGCACAGAGCATTGCGGATCTGGCCGCGGCCAACCTGAGCACCGAAGATTCAAAAAGTGCGCAGCTGATTAGCGCGGATAGCTCTGATGATGCGAGCGATAGCTCAGTGGAAAGCGTGGACGCGGCGAGCAGCGATGTTAGCGGCAGCAGCGTTGAAAGCGTGGATGTGAGCGGTAGCAGCCTGGAAAGCGTGGATGTGTCAGGCTCAAGCCTGGAAAGCGTGGATGATTCATCCGAAGATAGCGAAGAAGAAGAACTGCGCATTCTGAGCAGCAAAAAAAGCGGCAGCTATTATAGCTATGGTACGAAAAAAAGCGGCAGCTATAGCGGCTATAGCACCAAAAAATCGGCGAGCCGTCGCATCCTGAGCAGCAAAAAAAGCGGCTCGTATTCAGGCTATAGCACCAAAAAAAGCGGCAGCCGTCGCATTCTGAGCAGCAAAAAAAGCGGCTCATATTCTGGCAGCAAAGGCAGCAAACGCCGCATTCTGAGCAGCAAAAAAAGTGGCAGCTACAGCGGTAGCAAAGGCAGCAAACGCCGTAACCTGAGCAGCAAAAAAAGCGGTAGCTATAGCGGCTCGAAAGGCAGCAAACGTCGTATCCTGAGTAGCAAAAAAAGCGGCAGCTACTCGGGCAGCAAAGGCAGCAAACGTCGCAACCTGAGCAGCAAAAAAAGCGGCTCGTATAGCGGCAGTAAAGGCAGCAAACGTCGCATTCTGTCAGGCGGCCTGCGTGGCAGCATGTAA

image of the Sil1p protein structure without any modifications
This model on Alphafold demonstrates that this silaffin protein has a much greater structural confidence than the t.pseudonona silaffin protein did. this demonstrates that this diatom is probably more studied than t.pseudonona.
- Modify the Sil1p protein R5 peptide
R5 Peptide portion: SSKKSGSYSGSKGSKRRIL (from Claude and double checked through Googling the peptide)
Sil1p protein sequence R5 highlighted:
MKLTAIFPLLFTAVGYCAAQSIADLAAANLSTEDSKSAQLISADSSDDASDSSVESVDAASSDVSGSSVESVDVSGSSLESVDVSGSSLESVDDSSEDSEEEELRILSSKKSGSYYSYGTKKSGSYSGYSTKKSASRRILSSKKSGSYSGYSTKKSGSRRIL SSKKSGSYSGSKGSKRRIL SSKKSGSYSGSKGSKRRNL SSKKSGSYSGSKGSKRRIL SSKKSGSYSGSKGSKRRNL SSKKSGSYSGSKGSKRRIL SGGLRGSM
Modified Sil1p protein sequence with removal of R5 peptides:
MKLTAIFPLLFTAVGYCAAQSIADLAAANLSTEDSKSAQLISADSSDDASDSSVESVDAASSDVSGSSVESVDVSGSSLESVDVSGSSLESVDDSSEDSEEEELRILSSKKSGSYYSYGTKKSGSYSGYSTKKSASRRILSSKKSGSYSGYSTKKSGSRRILSSKKSGSYSGSKGSKRRNLSSKKSGSYSGSKGSKRRNLSGGLRGSM
reverse translation to a 624 base sequence of most likely codons.
atgaaactgaccgcgatttttccgctgctgtttaccgcggtgggctattgcgcggcgcagagcattgcggatctggcggcggcgaacctgagcaccgaagatagcaaaagcgcgcagctgattagcgcggatagcagcgatgatgcgagcgatagcagcgtggaaagcgtggatgcggcgagcagcgatgtgagcggcagcagcgtggaaagcgtggatgtgagcggcagcagcctggaaagcgtggatgtgagcggcagcagcctggaaagcgtggatgatagcagcgaagatagcgaagaagaagaactgcgcattctgagcagcaaaaaaagcggcagctattatagctatggcaccaaaaaaagcggcagctatagcggctatagcaccaaaaaaagcgcgagccgccgcattctgagcagcaaaaaaagcggcagctatagcggctatagcaccaaaaaaagcggcagccgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcaacctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcaacctgagcggcggcctgcgcggcagcatg
codon optimized for e.coli (https://en.vectorbuilder.com/tool/codon-optimization/cc081974-4c44-4101-975e-c3e8981e399b.html)
ATGAAACTGACCGCGATTTTTCCGCTGCTGTTTACCGCCGTGGGTTATTGCGCGGCCCAGTCTATTGCGGATCTGGCCGCGGCCAACCTGAGCACCGAAGATAGCAAAAGCGCGCAACTGATTAGCGCGGACAGCAGCGATGATGCGAGCGATAGCAGTGTGGAAAGCGTGGATGCCGCGAGCAGCGATGTGAGCGGCTCGAGCGTGGAAAGCGTGGATGTGAGCGGCAGCAGCCTGGAAAGTGTGGATGTGAGCGGCTCATCTCTGGAATCGGTGGATGATAGCAGCGAAGATTCCGAAGAAGAAGAACTGCGCATTCTGAGCAGCAAAAAAAGCGGCAGCTATTATAGCTATGGCACCAAAAAAAGCGGCAGCTATAGCGGCTATAGCACCAAAAAGAGCGCGTCGCGTCGCATTCTGTCAAGCAAAAAATCGGGCTCGTATTCAGGCTACAGCACCAAAAAGAGCGGCAGCCGCCGCATTCTGAGCAGCAAAAAAAGCGGCAGTTACAGCGGCAGCAAAGGCAGCAAACGTCGTAACCTGAGCAGCAAAAAAAGCGGCAGCTATAGCGGCAGCAAAGGCAGCAAACGCCGCAACCTGAGCGGCGGCCTGCGTGGCAGCATGTAA
Modified Sil1p protein sequence with overexpression of R5 peptides:
MKLTAIFPLLFTAVGYCAAQSIADLAAANLSTEDSKSAQLISADSSDDASDSSVESVDAASSDVSGSSVESVDVSGSSLESVDVSGSSLESVDDSSEDSEEEELRILSSKKSGSYYSYGTKKSGSYSGYSTKKSASRRILSSKKSGSYSGYSTKKSGSRRIL SSKKSGSYSGSKGSKRRIL SSKKSGSYSGSKGSKRRIL SSKKSGSYSGSKGSKRRNL SSKKSGSYSGSKGSKRRIL SSKKSGSYSGSKGSKRRIL SSKKSGSYSGSKGSKRRNL SSKKSGSYSGSKGSKRRIL SSKKSGSYSGSKGSKRRIL SGGLRGSM
MKLTAIFPLLFTAVGYCAAQSIADLAAANLSTEDSKSAQLISADSSDDASDSSVESVDAASSDVSGSSVESVDVSGSSLESVDVSGSSLESVDDSSEDSEEEELRILSSKKSGSYYSYGTKKSGSYSGYSTKKSASRRILSSKKSGSYSGYSTKKSGSRRILSSKKSGSYSGSKGSKRRILSSKKSGSYSGSKGSKRRILSSKKSGSYSGSKGSKRRNLSSKKSGSYSGSKGSKRRILSSKKSGSYSGSKGSKRRILSSKKSGSYSGSKGSKRRNLSSKKSGSYSGSKGSKRRILSSKKSGSYSGSKGSKRRILSGGLRGSM
reverse translation to a 966 base sequence of most likely codons.
atgaaactgaccgcgatttttccgctgctgtttaccgcggtgggctattgcgcggcgcagagcattgcggatctggcggcggcgaacctgagcaccgaagatagcaaaagcgcgcagctgattagcgcggatagcagcgatgatgcgagcgatagcagcgtggaaagcgtggatgcggcgagcagcgatgtgagcggcagcagcgtggaaagcgtggatgtgagcggcagcagcctggaaagcgtggatgtgagcggcagcagcctggaaagcgtggatgatagcagcgaagatagcgaagaagaagaactgcgcattctgagcagcaaaaaaagcggcagctattatagctatggcaccaaaaaaagcggcagctatagcggctatagcaccaaaaaaagcgcgagccgccgcattctgagcagcaaaaaaagcggcagctatagcggctatagcaccaaaaaaagcggcagccgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcaacctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcaacctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcagcaaaaaaagcggcagctatagcggcagcaaaggcagcaaacgccgcattctgagcggcggcctgcgcggcagcatg
codon optimized for e.coli (https://en.vectorbuilder.com/tool/codon-optimization/0bf0c7db-4609-4ca8-91c8-5f18fcb233bd.html)
ATGAAACTGACCGCGATTTTTCCGTTACTGTTTACCGCCGTCGGCTATTGTGCGGCGCAGAGCATTGCGGATCTGGCCGCGGCGAACCTGAGCACCGAAGATTCTAAAAGCGCGCAGCTGATTAGCGCGGATAGCAGCGATGATGCGAGCGATTCGAGCGTGGAAAGCGTAGATGCAGCCAGCTCTGATGTTAGCGGCAGCAGCGTGGAAAGCGTGGATGTTAGCGGCAGCAGCCTGGAAAGCGTGGATGTGAGCGGCAGCAGCCTGGAAAGCGTGGATGATAGCAGCGAAGATTCTGAAGAAGAAGAACTGCGGATTCTGAGCAGCAAAAAATCGGGCAGCTACTACAGCTATGGTACCAAAAAAAGCGGCTCATATAGCGGCTATAGCACCAAAAAAAGCGCGAGCCGTCGTATTCTGAGCAGCAAAAAAAGTGGCAGCTACAGCGGCTACAGCACCAAAAAAAGCGGCTCACGTCGTATTCTGAGCAGCAAAAAAAGCGGCAGCTACAGCGGCTCGAAAGGTAGCAAACGCCGCATTCTGTCAAGCAAAAAAAGCGGCAGCTACAGTGGTAGCAAAGGCAGCAAACGCCGCATCCTGAGCAGCAAAAAATCAGGCAGCTATAGCGGCAGCAAAGGCTCAAAACGTCGTAACTTAAGCAGCAAAAAATCGGGCAGCTACAGCGGCAGCAAAGGCAGCAAACGTCGTATCCTGTCGTCCAAAAAAAGCGGCAGCTATAGCGGCTCAAAAGGCAGCAAACGTCGCATCCTGAGCTCAAAAAAAAGCGGCAGCTATTCCGGCAGCAAAGGTTCCAAACGCCGCAACCTGAGCAGCAAAAAAAGCGGCTCTTATAGCGGCAGCAAAGGCAGCAAACGTCGCATTCTGAGCAGCAAAAAAAGCGGTAGCTATAGCGGCAGCAAAGGCAGCAAACGCCGCATTCTGAGCGGTGGTCTGCGCGGCAGCATGTAA
Other components to create the cell-free system
T7 promoter: TAATACGACTCACTATAGG
sources: https://www.neb.com/en-us/faqs/what-is-the-promoter-sequence-of-t7-rna-polymerase and screen shot of NEB. The screen shot had an extra G which I added to my benchling construct. Thank you Amanda for finding and screen shotting this cell free video for me!
RBS sequence: AGGAGG
Start Codone: ATG
Forward Primer: GCGAAT T7 promoter (TAATACGACTCACTATAGG) GCTTAAGTATA RBS(AGGAGG) AAAAAAT Start Codone (ATG)
Stop Codone: TAA
Other Base Pairs: NNNNNNNNNN
T7 Terminator: TAGCATAACCCCTTGGGGCCTCTAAACGGGTCTTGAGGGGTTTTTTG
Ending Sequence: TAANNNNNNNNNNTAGCATAACCCCTTGGGGCCTCTAAACGGGTCTTGAGGGGTTTTTTG
Detailed Workflow created through Claude
Step 1 — Codon Optimization and Construct Design (Week 1) Design three expression constructs encoding: (1) wild-type Sil1p (UniProt O74824), (2) Sil1p-ΔR5 with all five R5 repeat units removed, and (3) Sil1p-2xR5 with the R5 repeat array duplicated from 5 to 10 copies. All constructs include an N-terminal T7 promoter, Shine-Dalgarno sequence, His6-tag, and T7 terminator. Codon-optimize sequences for E. coli BL21 DE3 expression using Twist’s integrated codon optimization tool.
- Machine: None (computational design)
- Expected result: Three validated construct sequences ready for synthesis ordering
- Timeline: Days 1–3
Step 2 — Twist Bioscience DNA Order (Week 1) Submit all three constructs as whole plasmid synthesis orders to Twist Bioscience using the pTwist-T7 backbone. Select clonal gene synthesis for each construct.
- Machine: None (online order)
- Expected result: Sequence-verified plasmid DNA delivered within 7–10 business days
- Timeline: Days 3–4
Step 3 — Plasmid Receipt and Quality Check (Week 2) Upon receipt, resuspend lyophilized plasmid DNA per Twist instructions. Verify by Sanger sequencing and gel electrophoresis on a 1% agarose gel.
- Machine: ATC Thermal Cycler (for Sanger PCR), gel electrophoresis system
- Plate: 96-Armadillo-PCR-AB2396X for PCR setup
- Expected result: All three plasmids confirmed sequence-correct
- Timeline: Days 10–12
Step 4 — Cell-Free Reaction Setup (Week 2–3) Using the Echo525 acoustic liquid handler, transfer plasmid DNA (250 ng each) and Ginkgo BL21 DE3 cell-free master mix into a 384-well Echo PP plate in triplicate for each construct plus a no-template negative control. Total reaction volume: 5 µL per well.
- Machine: Echo525
- Plate: 384-well Echo PP
- Expected result: Consistent, low-volume transfers with <5% CV across replicates
- Timeline: Day 14
Step 5 — Plate Sealing and Cell-Free Expression (Week 3) Seal the 384-well plate with the Plateloc using A4s breathable seal to allow gas exchange during expression. Incubate in the Inheco Plate Incubator at 37°C for 4 hours.
- Machine: Plateloc, A4s seal, Inheco Plate Incubator
- Plate: 384-well Echo PP
- Expected result: Protein expression occurs in all construct wells; negative control shows no expression
- Timeline: Day 14, hours 0–4
Step 6 — TMOS Silica Precipitation Reaction (Week 3) Transfer 2 µL of each cell-free reaction to a fresh 384 Greiner black-well clear-bottom plate using the Bravo-384 plate stamp. Using the Multiflo dispenser, add 3 µL of freshly hydrolyzed tetramethyl orthosilicate (TMOS, 1M in 1mM HCl) to each well to initiate silica precipitation.
- Machine: Bravo-384, Multiflo
- Plate: 384 Greiner black-well clear-bottom
- Expected result: Visible silica precipitation in WT and 2xR5 wells within 5–10 minutes; reduced precipitation expected in ΔR5 wells
- Timeline: Day 14, hour 5
Step 7 — Mixing and Incubation (Week 3) Shake the precipitation plate on the BioshakeD3000 at 1,200 rpm for 20 minutes at room temperature to ensure complete silica polymerization.
- Machine: BioshakeD3000
- Plate: 384 Greiner black-well clear-bottom
- Expected result: Complete silica polymerization; visible white precipitate in active wells
- Timeline: Day 14, hours 5–5.5
Step 8 — Centrifugation to Pellet Silica (Week 3) Centrifuge the precipitation plate at 3,000 × g for 10 minutes in the HiG Centrifuge to pellet silica particles.
- Machine: HiG Centrifuge
- Plate: 384 Greiner black-well clear-bottom
- Expected result: Silica pellet visible at well bottom; clear supernatant containing residual free silicic acid
- Timeline: Day 14, hour 6
Step 9 — Supernatant Transfer for Colorimetric Assay (Week 3) Use the Bravo-384 to transfer 4 µL of supernatant from each well into a fresh 384-flat-corning-3640 plate for silicomolybdate assay. Retain the pellet plate for spectrum scanning in Step 11.
- Machine: Bravo-384
- Plate: 384-flat-corning-3640
- Expected result: Clean supernatant transfer without disturbing silica pellets
- Timeline: Day 14, hour 6.5
Step 10 — Silicomolybdate Colorimetric Assay (Week 3) Using the Tempest bulk dispenser, add 1 µL of silicomolybdate reagent (ammonium molybdate in sulfuric acid) to each supernatant well. Incubate 10 minutes at room temperature. Read absorbance at 810 nm on the Spark Plate Reader. Lower absorbance in the supernatant = more silica precipitated by the protein.
- Machine: Tempest, Spark Plate Reader
- Plate: 384-flat-corning-3640
- Expected result: WT and 2xR5 wells show lower A810 than ΔR5 and no-template control, indicating greater silica precipitation
- Timeline: Day 14, hours 7–8
Step 11 — Full Spectrum Absorbance Scan of Silica Pellets (Week 3) Resuspend silica pellets in 5 µL ultrapure water by pipetting. Run a full-spectrum absorbance scan from 400–800 nm on the Spark Plate Reader to capture any optical differences between silica nanoparticles produced by different Sil1p variants.
- Machine: Spark Plate Reader
- Plate: 384 Greiner black-well clear-bottom
- Expected result: Spectral differences between WT, ΔR5, and 2xR5 silica particles; potential blue shift in 2xR5 particles if larger particle size shifts photonic scattering
- Timeline: Day 14, hour 8.5
Step 12 — Expression Confirmation: SDS-PAGE (Validation A, Week 3) Collect 5 µL of cell-free reaction from each construct well before TMOS addition. Load onto SDS-PAGE gel alongside a His-tag protein ladder. Run at 200V for 35 minutes. Stain with Coomassie blue and image.
- Machine: SDS-PAGE system (manual)
- Expected result: Bands at expected molecular weights: WT Sil1p (~21 kDa), Sil1p-ΔR5 (~14 kDa), Sil1p-2xR5 (~30 kDa)
- Timeline: Day 15
Step 13 — qPCR Expression Verification (Week 3) As an orthogonal expression check, extract total RNA from cell-free reactions and run qPCR using primers flanking the His6-tag sequence to confirm transcript levels across all three constructs are comparable.
- Machine: CFX Opus qPCR machine
- Plate: 96-Armadillo-PCR-AB2396X
- Expected result: Similar Ct values across all three constructs, confirming equivalent transcription; any differences in protein yield are post-transcriptional
- Timeline: Day 15
Step 14 — Data Analysis and Construct Ranking (Week 4) Compile colorimetric assay data, full-spectrum scans, SDS-PAGE results, and qPCR data. Calculate silica precipitation efficiency for each construct as: % silica precipitated = (A810 no-template − A810 construct) / A810 no-template × 100. Rank constructs by precipitation efficiency and spectral shift magnitude.
- Machine: None (computational analysis)
- Expected result: Clear ranking of Sil1p-2xR5 > WT Sil1p > Sil1p-ΔR5 in silica precipitation efficiency
- Timeline: Days 16–18
Step 15 — Iteration and Construct Refinement (Week 4–5) Based on results, design a second round of constructs if needed — e.g., Sil1p-3xR5, Sil1p with randomized repeat spacing, or Sil1p with non-native repeat sequences. Order from Twist Bioscience and re-enter the cell-free pipeline. This iteration loop establishes the quantitative R5 dose-response relationship that forms the foundation for Aim 2 diatom expression work.
- Machine: Echo525, Inheco, Spark (full pipeline repeat)
- Expected result: Refined structure-function map of R5 repeat domains
- Timeline: Days 18–25
Results and Quantitative Expectations


Benchling
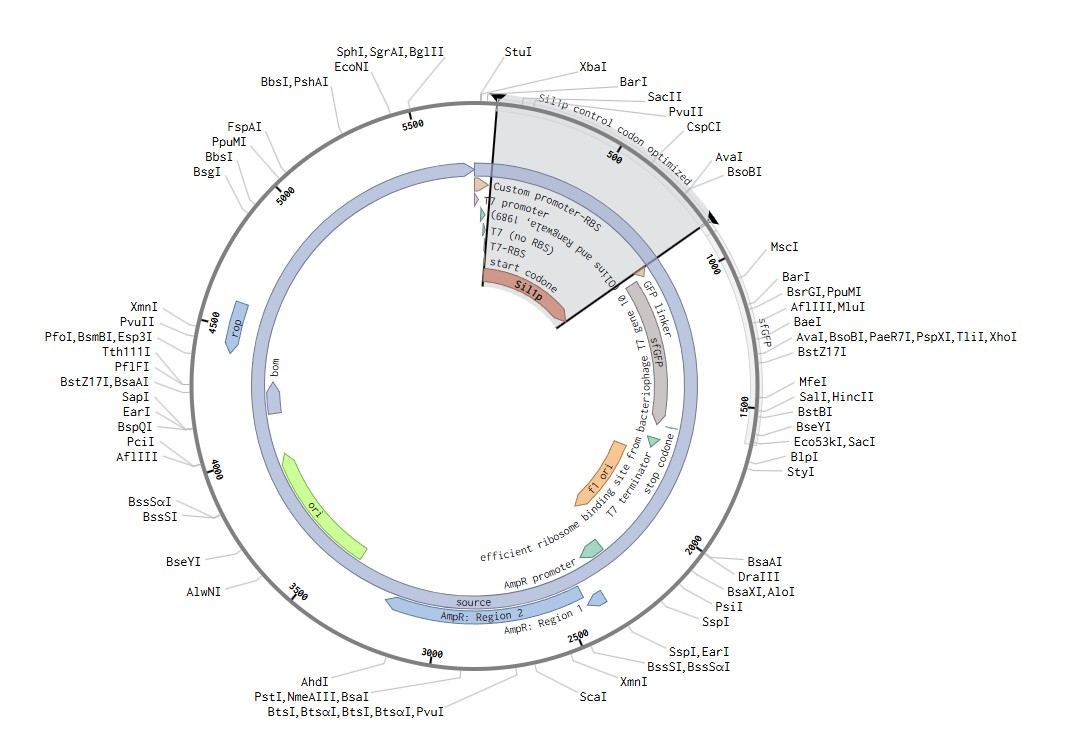
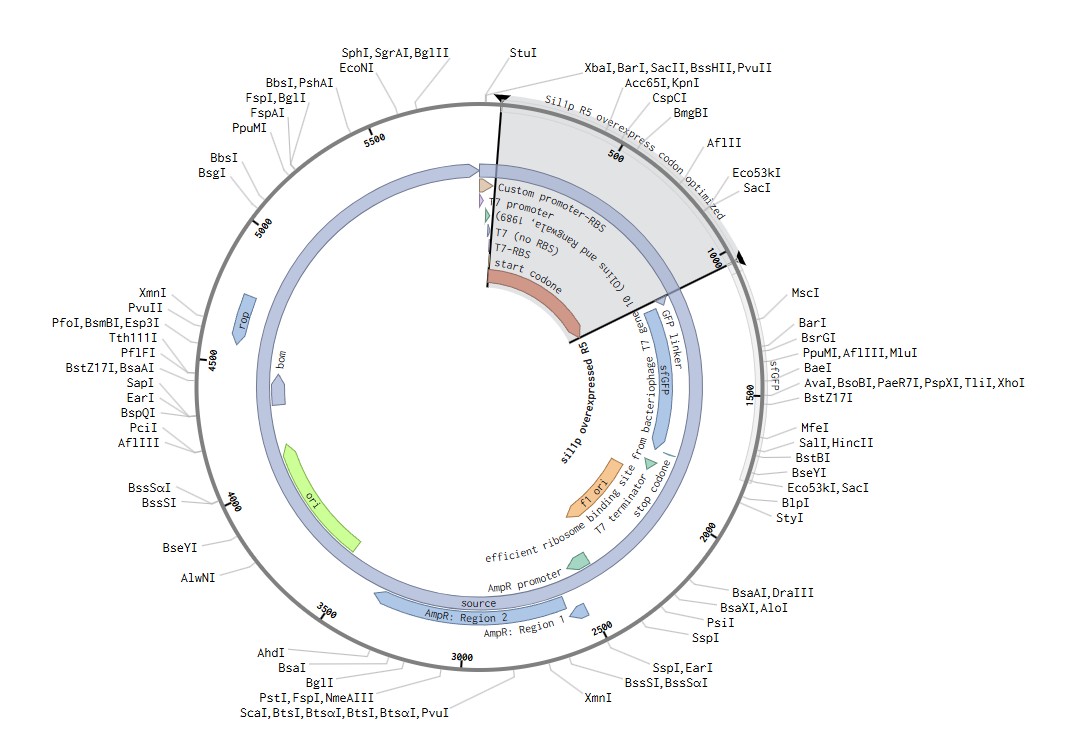
These cell-free constructs were created to run in the automated lab at Ginkgo. This means that it was created following the NEBExpress protocol, the same one noted in the cell-free lab and one that Ginkgo is familiar with, just with my constructs as the modifier for the experiment.
Technique Checklist (with Claude support)
- DNA design and synthesis (Twist Bioscience plasmid synthesis)
- Cell-free protein expression
- Colorimetric enzymatic assay (silicomolybdate / molybdenum blue)
- Full-spectrum absorbance spectroscopy
- Acoustic liquid handling (Echo525)
- Automated plate handling and sealing (Plateloc, XPeel, Bravo-384)
- High-throughput incubation (Inheco Plate Incubator, BioshakeD3000)
- SDS-PAGE protein gel electrophoresis
- qPCR gene expression quantification (CFX Opus)
- Microplate reader assay development (Spark)
- Bioinformatics / codon optimization
- GenBank construct annotation
The constructs are tagged with sfGFP to floresce if the synthesis was successful, but a colorimetric assay with the spark plate reader would be the best way to see the variation in light scattering that the three different constructs will demonstrate. Since color is a physical structural condition for diatoms, the three modified silica deposit constructs are expected to express color differently and demonstrate a different spectral shift.
These two tests will result in a table that notes whether the sample floresced, and if so, what the spectral reading comes out with. I expect a reading with some green and red, resulting in a brown color, however because it’s in a cell-free system, it may be different since the other parts of the cell system are not present in the same way, however that base variation should be consistent across all three constructs, so the variation is still quantifiable, but may not be representative of the final color expression for Aim 3 as the modified proteins are re-introduced into the diatoms.
This summer, Ronan and I plan to conduct the Aim 1 experiment! I am excited to learn more about the automated lab system and to see if my hypothesis unfolds or if the protein modifications do something unexpected.
Additional Information
For the Sil1p R5 peptide info
• Poulsen, N., Chesley, P.M. & Kröger, N. (2006). Molecular Genetic Manipulation of the Diatom Thalassiosira pseudonana. Journal of Phycology, 42, 1059–1065.
• Tesson B, Lerch SJL, Hildebrand M. Characterization of a New Protein Family Associated With the Silica Deposition Vesicle Membrane Enables Genetic Manipulation of Diatom Silica. Sci Rep. 2017 Oct 18;7(1):13457. doi: 10.1038/s41598-017-13613-8. PMID: 29044150; PMCID: PMC5647440.
Other diatom engineering sources
• Serif, M., Dubois, G., Finoux, AL. et al. One-step generation of multiple gene knock-outs in the diatom Phaeodactylum tricornutum by DNA-free genome editing. Nat Commun 9, 3924 (2018). https://doi.org/10.1038/s41467-018-06378-9
• Li Y, Deng L, Walker EJL, Karas BJ, Mock T. Genetic engineering in diatoms: advances and prospects. Plant J. 2025 Mar;121(6):e70102. doi: 10.1111/tpj.70102. PMID: 40089910; PMCID: PMC11910954.
• Tong, C.Y., Derek, C.J.C. The role of substrates towards marine diatom Cylindrotheca fusiformis adhesion and biofilm development. J Appl Phycol 33, 2845–2862 (2021). https://doi.org/10.1007/s10811-021-02504-1
Budget
Assuming that the lab (in this case, Ginkgo Bioworks), has the plate reader and other equipment needed to conduct the cell-free automated experiment, the price of the Aim 1 experiment is limited to the cost of the Twist order. The cost is fairly high, and for that, Ronan may have only ordered two of the constructs to best balance the cost and time per student on his end at Ginkgo.
Alternative project approaches
Another potential project was to create a touch-reactive biofilm that pigmented with pressure. This could use a system along the lines of the cell-free systems that were discussed in the Week 8 lecture. However for this project, I wanted the color to fade away once the pressure was lessened, however that may not be possible with cell-free systems as they seem to be a one-time reaction.
Alternatively to diatoms, it could also be interesting to use mycelium and genetically modify it to be clear. This uses the strength and potential for mycelium as a building material, binding to architectural waste like glass pieces to create a composite material with the tensile strength and growth rate of mycelium. A clear mycelium would entail a melanin gene knockout, and potentially doing so with a more clear mycelium strain to begin with. This method was developed with the help of CRISPR.
Claude generated information:
SECTION 5: TECHNIQUES, TOOLS, AND TECHNOLOGY
Technique Expansion
1. Cell-Free Protein Synthesis (CFPS)
Cell-free protein synthesis (CFPS) is an in vitro method for producing proteins directly from DNA templates without the use of living cells. The system consists of a cell lysate — in this project, BL21 DE3 lysate prepared at Ginkgo Bioworks — combined with a master mix containing ribosomes, amino acids, energy regeneration components, and RNA polymerase. When a plasmid encoding a T7 promoter-driven gene is added, T7 RNA polymerase transcribes the gene into mRNA, which is then translated by ribosomes present in the lysate into protein. CFPS is particularly powerful for this project because it allows multiple silaffin variants to be expressed and assayed in parallel in a 384-well format within a single day, without the need for bacterial transformation, overnight culture, or IPTG induction — dramatically accelerating the design-build-test cycle for protein engineering.
2. Silicomolybdate Colorimetric Assay
The silicomolybdate assay, also known as the molybdenum blue assay, is a well-established colorimetric method for quantifying free silicic acid (Si(OH)₄) in solution. In the presence of ammonium molybdate under acidic conditions, free silicic acid forms a yellow silicomolybdate complex; upon reduction with ascorbic acid or other reducing agents, this complex turns an intense blue color with peak absorbance at 810 nm. In this project, the assay is applied to the supernatant after silica precipitation — the more silica the Sil1p variant has precipitated from solution, the less free silicic acid remains, and therefore the lower the A810 reading. This indirect measurement elegantly reports on the silica-precipitating activity of each Sil1p variant in a format fully compatible with automated 384-well plate reading on the Spark platform, enabling quantitative comparison of precipitation efficiency across all three constructs simultaneously.
SECTION 6: PROJECT VALIDATION
10a — Validation Choice
Two complementary validation experiments are planned for this project, to be performed based on available lab access and timeline. Validation A (SDS-PAGE) directly confirms that all three Sil1p variants are being produced as proteins of the expected size in the cell-free system, ruling out expression failure as a confound. Validation B (silicomolybdate pilot assay) directly confirms that the wild-type Sil1p is functionally active in precipitating silica from TMOS, and that the ΔR5 truncation reduces this activity — establishing the functional assay and the expected directionality of results before the full 384-well campaign is run.
10b — Validation Protocols
Validation A: SDS-PAGE Expression Confirmation
- Collect 5 µL of cell-free reaction from each of the three Sil1p constructs and the no-template control after 4 hours of expression.
- Add 5 µL of 2× Laemmli SDS sample buffer to each sample.
- Heat samples at 95°C for 5 minutes using the ATC Thermal Cycler.
- Load 8 µL of each sample onto a 4–20% gradient SDS-PAGE gel alongside a His-tag protein molecular weight ladder.
- Run electrophoresis at 200V for 35 minutes in Tris-glycine SDS running buffer.
- Stain gel with InstantBlue Coomassie stain for 30 minutes.
- Destain in ultrapure water for 20 minutes.
- Image gel using a gel documentation system.
- Confirm bands at expected molecular weights: WT Sil1p ~21 kDa, Sil1p-ΔR5 ~14 kDa, Sil1p-2xR5 ~30 kDa.
- Confirm absence of bands in no-template control lane.
Validation B: Silicomolybdate Pilot Assay
- Set up cell-free reactions for WT Sil1p, Sil1p-ΔR5, and no-template control in triplicate in a 96-well plate (25 µL reactions).
- Incubate at 37°C for 4 hours in the Inheco Plate Incubator.
- Add 5 µL of freshly hydrolyzed TMOS (1M in 1mM HCl) to each well.
- Shake on BioshakeD3000 at 1,200 rpm for 20 minutes at room temperature.
- Centrifuge at 3,000 × g for 10 minutes in the HiG Centrifuge to pellet silica.
- Transfer 20 µL of supernatant to a fresh 96-well flat-bottom plate.
- Add 5 µL of silicomolybdate reagent (0.026M ammonium molybdate in 0.1M H₂SO₄) to each well.
- Incubate 10 minutes at room temperature.
- Add 5 µL of reducing solution (0.1M ascorbic acid) if molybdenum blue endpoint is desired.
- Read absorbance at 810 nm on the Spark Plate Reader.
- Calculate % silica precipitated relative to no-template control.
- Confirm WT Sil1p shows significantly lower A810 than ΔR5 and no-template control (expected: >40% reduction).
10c — Techniques Used
SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis) separates proteins by molecular weight under denaturing conditions, allowing direct visualization of whether each Sil1p variant has been produced at the correct size and in sufficient quantity by the cell-free system. The silicomolybdate assay is a spectrophotometric technique that quantifies free silicic acid in solution through formation of a chromogenic molybdate complex, providing an indirect but highly sensitive measure of silica precipitation activity. Cell-free protein synthesis leverages the transcription and translation machinery of bacterial lysates to produce proteins from plasmid DNA templates in vitro, enabling rapid, parallel expression of multiple variants without live organism handling. Together, these three techniques — SDS-PAGE, colorimetric spectrophotometry, and cell-free expression — form an integrated validation pipeline that confirms both the production and the functional activity of each engineered Sil1p variant before proceeding to the full high-throughput 384-well campaign.
Troubleshooting
The most likely technical challenge is low or absent silaffin protein expression in the cell-free system, which could result from poor codon optimization, mRNA secondary structure inhibiting translation, or issues with the His6-tag affecting protein folding. If expression is not confirmed by SDS-PAGE, the first corrective step would be to re-optimize the 5’ UTR sequence using the Salis Lab RBS Calculator and reorder a revised construct from Twist Bioscience before repeating the cell-free expression. A second potential challenge is non-specific silica precipitation in the no-template control wells — TMOS is chemically reactive and will self-polymerize under certain pH conditions independently of silaffin; this can be mitigated by carefully controlling TMOS hydrolysis conditions (1mM HCl, room temperature, freshly prepared) and including a TMOS-only control well in every plate. A third limitation is that cell-free silica precipitation may not faithfully recapitulate the in vivo post-translational modifications of native Sil1p — including phosphorylation and polyamine modifications on lysine residues — which are known to enhance biosilicification activity; this is an inherent constraint of the cell-free platform that will need to be addressed in Aim 2 when the work moves into a native diatom expression system. Finally, the full-spectrum scan for structural color differences is exploratory and may not yield interpretable optical signals at the scale of cell-free silica nanoparticles — if this is the case, dynamic light scattering (DLS) would be pursued as an alternative to characterize particle size distributions.
Claude References
- Kröger, N., Deutzmann, R., & Sumper, M. (1999). Polycationic peptides from diatom biosilica that direct silica nanosphere formation. Science, 286(5442), 1129–1132.
- Sumper, M., & Brunner, E. (2008). Silica biomineralisation in diatoms: the model organism Thalassiosira pseudonana. ChemBioChem, 9(8), 1187–1194.
- Kröger, N., Lorenz, S., Brunner, E., & Sumper, M. (2002). Self-assembly of highly phosphorylated silaffins and their function in biosilica morphogenesis. Science, 298(5593), 584–586.
- Poulsen, N., Sumper, M., & Kröger, N. (2003). Biosilica formation in diatoms: characterization of native silaffin-2 and its role in silica morphogenesis. PNAS, 100(21), 12075–12080.
- Lechner, C. C., & Becker, C. F. (2015). Silaffins in silica biomineralization and biomimetic silica precipitation. Marine Drugs, 13(8), 5297–5333.
- Twist Bioscience. (2024). Whole plasmid synthesis ordering guide. https://www.twistbioscience.com/products/genes
- Ginkgo Bioworks. (2024). Cell-free expression platform. https://www.ginkgobioworks.com
- Opentrons. (2024). OT-2 liquid handling robot. https://opentrons.com
- SecureDNA. (2024). DNA synthesis screening. https://securedna.org