Individual Final Project: A Self-Lytic E. coli Chassis for Cost-Effective Biopolymer Production via Rational Circuit Design
SECTION 1: ABSTRACT
Plastic pollution is one of the biggest challenges of 21st century: We have around 7 gr of microplastics in our brains ( Nihart, A.J., Garcia, M.A., El Hayek, E. et al. Bioaccumulation of microplastics in decedent human brains. Nat Med 31, 1114–1119, 2025). Using microorganisms it is already possible to produce biodegradable plastics that do not accumulate in our bodies.
The transition from petroleum-based plastics to sustainable biopolymers, such as polyhydroxyalkanoates (PHAs), is hindered by prohibitive production costs. While metabolic engineering has significantly improved cellular yields, downstream processing (DSP)—specifically cell disruption—remains a critical economic bottleneck, accounting for 30–40% of total operating expenses. Current research into continuous secretion systems shows promise for small molecules; however, biopolymers like PHAs form large, insoluble intracellular granules that are biophysically difficult to export without excessive metabolic cost.
This project proposes a “Self-Lytic” chassis that utilizes programmed autolysis to bypass these limitations. We hypothesize that a dual-input AND-gate logic circuit, integrating phosphate-starvation (pPhoA) and temperature-induction (thermo-riboswitch) will provide precise control over cell lysis, facilitating rapid and cost-effective PHA recovery.
To optimize this circuit, we will employ a computational rational design approach. We will use the Salis Lab RBS Calculator to predict translation initiation rates and stochastic simulation + absolute quantification to simulate the threshold of holin accumulation required for lysis. During the pre-lysis growth and polymer accumulation phase, Flux Balance Analysis (FBA) will be used to model the metabolic burden imposed by circuit maintenance on the host, ensuring that expression of the biosynthetic and regulatory components does not unsustainably compete with PHA synthesis for cellular resources. This predictive framework replaces high-throughput physical screening, allowing us to select a curated library of specific RBS variants that minimize basal leakiness and maximize polymer yield.
By shifting from energy-intensive mechanical homogenization to programmed biological lysis, this platform aims to reduce downstream costs by up to 60%, offering a scalable, open-source solution for sustainable biomanufacturing.
SECTION 2: PROJECT AIMS
Aim 1: Experimental Aim (this project): The first aim of my final project is to engineer and characterize a dual-input (phosphate-starvation and temperature-inducible) genetic AND-gate for programmed autolysis in E. coli by utilizing computational thermodynamic modeling and kinetic simulations. Specifically, I will use the Salis Lab RBS Calculator to design a curated library of 5-10 Ribosome Binding Sites with predicted translation initiation rates to control the expression of the Lambda phage lysis cassette (SRRz). The circuit’s performance will be modeled using stochastic simulation in Python to predict the lysis threshold. Flux Balance Analysis (FBA) will complement this by modeling the metabolic burden imposed by basal circuit expression on the host during the pre-lysis growth and polymer accumulation phase. Experimental validation will involve cloning the synthetic circuit via NEB Gibson Assembly and characterizing lysis kinetics through OD600 absorbance assays. Nile Red fluorescence microscopy will be used to confirm intracellular PHA granule accumulation prior to lysis induction and to verify granule release upon lytic activation.
Aim 2: Development Aim: The next step following a successful Aim 1 is to solve the “leakiness” and scalability challenges in high-density industrial fermentations by implementing a machine learning model that will be trained on fermentation process variables — including dissolved oxygen, pH, osmolarity, and nutrient consumption rates — collected across scales, with the goal of predicting optimal lysis induction timing as a function of bioreactor conditions. This data-driven layer will serve as a process control strategy to ensure that lytic activation remains synchronized with peak polymer accumulation regardless of scale-dependent environmental heterogeneity. This aim focuses on transitioning the lytic circuit from lab-scale E. coli to testing the circuit’s robustness in 30L, 100L, and 1000L bioreactors to ensure that the lysis trigger remains perfectly synchronized with peak PHA accumulation under the fluctuating nutrient gradients and high osmotic pressures typical of industrial scales.
Aim 3: Visionary Aim: The long-term vision for this project is to establish an open-source, “Self-Harvesting” platform that disrupts the current economic paradigms of intracellular biomanufacturing. By eliminating the need for capital-intensive mechanical cell disruption and reducing dependency on toxic chemical solvents, this technology aims to bring the price of bioplastics to parity with petroleum-based polymers. Beyond PHAs, this project envisions a collaborative ecosystem where researchers can share experimental results, refine computational models, and contribute new genetic parts. This open-source framework will not be limited to E. coli; it is designed for chassis-portability, allowing the “self-lytic” logic to be adapted to diverse organisms such as Halomonas, or cyanobacteria. Ultimately, this platform enables a decentralized circular bioeconomy, democratizing the production of single-cell proteins, enzymes, and specialty chemicals, and fostering a global community dedicated to sustainable, high-value biological manufacturing.
SECTION 3: BACKGROUND Two key research citations inform this project:
Holin-Endolysin Systems: Research by Young et al. (2000) characterizes the Lambda phage SRRz lytic core, demonstrating that holins form micron-scale pores in the inner membrane while endolysins degrade the peptidoglycan layer. This two-component system provides the biological “explosive” necessary for rapid cell disruption.
Phosphate-Starvation Induction: Studies on the pho regulon (pPhoA promoter) show that E. coli can be programmed to respond to inorganic phosphate depletion, a condition that naturally coincides with the onset of PHA accumulation. This suggests that the signal for polymer production can be dual-purposed as the trigger for harvesting, ensuring metabolic synchronization.
Innovation and Novelty: This project departs from the traditional “brute force” paradigm of mechanical lysis, adopting a predictive synthetic biology approach to downstream processing. While holin-endolysin systems have been explored in E. coli, they typically rely on single-input induction, leaving them susceptible to premature activation under fluctuating industrial conditions.
The innovation lies in the integration of a dual-input AND-gate logic circuit (pPhoA and thermal induction), which provides a level of conditional specificity currently absent in PHA recovery strategies. By utilizing thermodynamic modeling (Salis Lab RBS Calculator) and kinetic ODE simulations, this design enables the rational selection of Ribosome Binding Sites (RBS) to precisely define the holin accumulation threshold. This predictive framework constrains the translation initiation space, effectively minimizing the basal leakiness that typically plagues autolytic strains. Ultimately, this transforms the cellular chassis into a programmable, self-disrupting unit, expanding the boundaries of synthetic biology into the realm of autonomous downstream processing.
Significance and Impact: This project addresses the global crisis of plastic pollution by tackling the high cost of sustainable alternatives. The economic bottleneck of downstream processing represents a critical barrier that keeps bioplastics from competing with cheap, petroleum-derived synthetics. Reducing harvesting costs could catalyze a field-level change, shifting the industry toward biological, energy-efficient extraction. Beyond the immediate research, the outcomes of this project contribute to a broader societal transition toward a circular bioeconomy, reducing our carbon footprint and reliance on fossil fuels. If realized, this open-source framework will democratize biomanufacturing, enabling the decentralized production of high-value compounds in resource-limited settings and fostering global technical collaboration.
Ethical Implications: By developing an open-source platform for cheaper bioplastics, the project promotes the principle of beneficence, aiming to mitigate the environmental damage caused by traditional plastics. However, the use of engineered “self-destructive” bacteria raises concerns regarding Biosecurity and Responsibility. There is a potential risk of accidental release of engineered strains into the environment; although the autolytic circuit acts as a built-in “kill-switch” outside of controlled conditions, the horizontal gene transfer of antibiotic resistance markers (if used) remains an ethical challenge that must be addressed to ensure environmental safety.
To ensure the project is conducted ethically, I propose the implementation of “Safe-by-Design” biocontainment strategies, such as using auxotrophic strains that cannot survive outside the laboratory or replacing antibiotic markers with metabolic complementation. Potential unintended consequences include the misuse of the autolytic logic to engineer pathogens, which necessitates a commitment to the principle of Responsibility through transparent, but regulated, sharing of genetic parts. We must also acknowledge the uncertainty in our assumptions regarding the scalability of lysis; if the circuit fails to trigger in large bioreactors, it could lead to significant waste of resources. Alternatives to this action include chemical surfactants, but these are less sustainable and pose higher toxicological risks to workers and the environment. SECTION 4: EXPERIMENTAL DESIGN, TECHNIQUES, TOOLS, AND TECHNOLOGY Use Claude AI skills to refine your HTGAA final project experimental design here
Section 4: Experimental Design
4.1 Workflow Overview
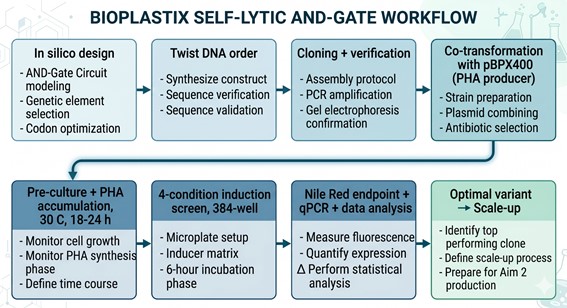
4.2 Detailed Step-by-Step Protocol (15 steps)
The following 15-step workflow covers in silico circuit design, Twist synthesis, strain construction, dual-input induction, lysis characterization, PHA quantification, RBS library optimization, FBA validation, and data integration.
| Step | Method or Tool | Equipment | Plate type | Expected result | Timeline |
|---|---|---|---|---|---|
| 1. Computational RBS design | Salis Lab RBS Calculator (web) + stochastic ODE simulation (scipy.integrate.solve_ivp); FBA burden via cobrapy on iML1515 | Desktop (Python) | — | 5 RBS sequences with predicted TIR ~200, 1.5k, 10k, 50k, 200k a.u. | 1 Week |
| 2. Twist Bioscience order | 1 × Custom Plasmid (pBioplastix-Lysis, 5,829 bp, verified pSC101 ori) + 5 × eBlock Gene Fragments (~390 bp each); SecureDNA pre-screen | Online order portal | - | DNA delivered as lyophilized stocks | 14–18 days (lead time) |
| 3. Backbone linearization | Q5 inverse PCR around S105 RBS site to drop the wild-type RBS region | ATC Thermal Cycler | 96-Armadillo-PCR-AB2396X | Linear ~5.5 kb backbone, single band on E-gel | 4 h |
| 4. Gibson assembly setup | Echo525 acoustic transfer of linearized backbone (50 ng) + each eBlock variant (3:1 molar excess) + NEBuilder HiFi master mix (nL volumes); 5 reactions | Echo525 Acoustic Liquid Handler | 384-well Plate Echo PP (source) → 384-pcr-eppendorf-9510207XX (rxn) | 5 Gibson reactions, ~5 µL each | 30 min |
| 5. Gibson reaction | 50 °C × 15 min isothermal | ATC Thermal Cycler | 384-pcr-eppendorf-9510207XX | Assembled circular plasmids | 30 min |
| 6. Heat-shock transformation + outgrowth | Manual transformation of NEB 5-alpha competent cells; 1 h SOC outgrowth at 37 °C; plate on LB+Kan50 | Inheco Plate Incubator (outgrowth); manual plating | 96-Armadillo-PCR-AB2396X (outgrowth); 1-flat-thermo-264728-omni-96 (selection) | ~50–500 colonies/variant | 1 day |
| 7. Colony PCR + Sanger verification | Echo525 dispenses Q5 master mix + variant-specific primers; ATC thermal cycler; outsourced Sanger of positives | Echo525 + ATC | 384-pcr-eppendorf-9510207XX | 3 verified colonies/variant | 2–3 days |
| 8. Glycerol stock banking | 96-deep-well overnights (LB+Kan, 30 °C); Bravo-96 stamp into glycerol stock plates (25% v/v) | Cytomat (incubation); Bravo-96 (stamp) | 96-v-eppendorf-951033502-deep (overnight); 96-round-axygen-pdw11cs-halfdeep (stocks) | Banked at –80 °C; working stocks in Tundrastore (4 °C) | 1 day |
| 9. Co-transformation with pBPX400 | Heat-shock co-transformation into BL21(DE3); dual selection LB+Kan50+Cm25 | Inheco Plate Incubator; manual plating | 1-flat-thermo-264728-omni-96 | Dual-resistant strains: 5 × pBioplastix-Lysis(V1–V5) + pBPX400 | 1 day |
| 10. Pre-culture + PHA accumulation | Inoculate from glycerol stocks into PHA-accumulation MOPS minimal media (0.4% glucose + 4 mM Pi, 30 °C); 18–24 h | Bravo-96 (inoculation); Cytomat (shaking incubation, 30 °C, 600 rpm) | 96-v-eppendorf-951033502-deep | OD600 ≈ 3–5; intracellular PHB granules visible by Nile Red microscopy | 18–24 h |
| 11. Transfer to assay plate + induction setup | Echo525 transfers 1:100 dilutions into 384-well assay plate; Multiflo bulk-dispenses induction media (low-Pi MOPS for ON, high-Pi MOPS for OFF) | Echo525; Multiflo Automated Microplate Dispenser | 384 Greiner black-well clear-bottom (40 µL/well) | Assay plate ready: 5 variants × 4 conditions × 3 reps + controls | 30 min |
| 12. Induction + kinetic readout | Plates split: OFF/OFF + Pi-only conditions stay in Cytomat at 30 °C; heat-only + ON/ON move to Inheco at 42 °C; Spark Plate Reader kinetic reads every 10 min × 6 h (OD600 + sfGFP Ex 485/Em 520) | Cytomat (30 °C); Inheco (42 °C); Spark Plate Reader | 384 Greiner black-well clear-bottom; sealed with Plateloc + A4s breathable | OD600 trajectories + sfGFP induction kinetics | 6 h |
| 13. Nile Red endpoint assay | Tempest dispenses Nile Red (10 µg/mL final); BioshakeD3000 mix 30 min; HiG centrifuge to pellet debris; PHERAstar FSX reads supernatant fluorescence (Ex 543 / Em 598) | Tempest; BioshakeD3000; HiG Centrifuge; PHERAstar FSX | Same 384 Greiner plate | Nile Red supernatant fluorescence ~ released PHA mass | 1 h |
| 14. qRT-PCR transcript validation | Total RNA extraction (3 selected variants × 4 conditions × 3 reps, n=36); reverse transcription + qPCR for sfgfp, S, R, tetR, and rrsA (reference) | CFX Opus qPCR | 96-Armadillo-PCR-AB2396X | Confirms transcriptional AND-gate logic; validates anti-TetR depletion | 2 days |
| 15. Data analysis + variant ranking | Python pipeline: lysis kinetic parameter extraction (scipy.optimize.curve_fit on logistic decay); leakiness scoring; Pareto-front ranking; correlation of measured kinetics with predicted TIR | Desktop (Python, pandas, matplotlib) | — | Optimal RBS variant identified for Aim 2 | 1 week |
Conclusion: Total project duration: ~6–8 weeks (DNA lead time dominates).
4.3 Plate Layout — The 384-Well Phenotype Screen (Step 12)
The screening plate carries 5 RBS variants (V1–V5) and 5 control strains across 4 induction conditions, in 3 biological replicates. Edge wells (column 1, column 24, row A, row P) are filled with sterile media as an evaporation moat.
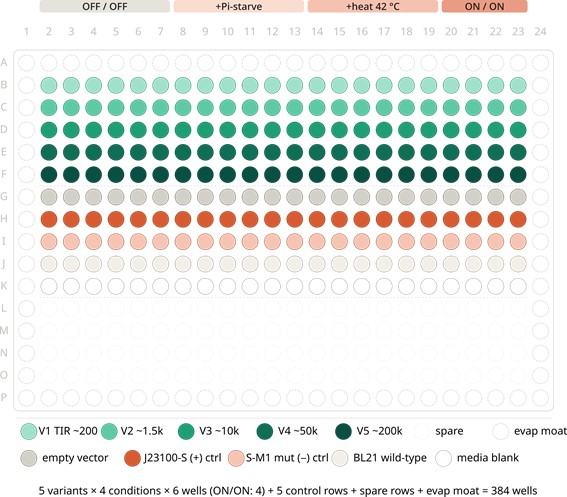
4.4 Standard Curves and Controls
The plate also includes a PHA standard curve: pure crotonic acid (PHB hydrolysate) standards at 0, 1, 5, 10, 25, 50 µg/mL spiked into Nile Red working solution (rows L–O, cols 2–7), allowing absolute quantification of released polymer mass per well. Nile Red fluorescence-vs-concentration is fit to a 4-parameter logistic curve, and sample wells are interpolated against this fit.
4.5 Twist Bioscience Order — Sequences
4.5.1 Base Plasmid (Twist Custom Plasmid order)
The full 5,829 bp pBioplastix-Lysis is ordered as a single Twist Custom Plasmid in the verified pSC101 / KanR backbone. Key features (positions referenced to GenBank file pBioplastix-Lysis.gb):

4.5.2 RBS Variant eBlocks (5 × Twist Gene Fragments, ~390 bp each) Each eBlock contains a 30 bp 5′ Gibson homology arm matching the end of sfGFP, the variant RBS (12 bp), the full 321 bp S105 ORF, and a 30 bp 3′ Gibson homology arm matching the R-RBS region. The only sequence change between eBlocks is the 12 bp RBS region (uppercase below); all flanking sequence is identical to the base plasmid.

Variant RBS sequences (output from Salis Lab RBS Calculator; values are illustrative — replace with calculator output before order)
| Variant | RBS sequence (12 bp) | Predicted TIR (a.u.) | log10(TIR) |
|---|---|---|---|
| V1 | aattcaaccaaa | ~200 | 2.3 |
| V2 | aaaggaagaaaa | ~1,500 | 3.2 |
| V3 | aaaggaggaata | ~10,000 | 4.0 |
| V4 | aaagaggagaaa | ~50,000 | 4.7 |
| V5 | aaaaggaggtaa | ~200,000 | 5.3 |
Section 5: Techniques, Tools, and Technology
5.1 Course Technique Checklist
| Technique | Used in this project? | Where in workflow |
|---|---|---|
| Gibson Assembly (NEBuilder HiFi) | ✓ | Step 4–5 (variant installation) |
| Computational RBS design (Salis Lab Calculator) | ✓ | Step 1 |
| Stochastic ODE simulation | ✓ | Step 1 |
| Flux Balance Analysis (FBA) | ✓ | Step 1 (burden assessment) |
| Polymerase chain reaction (PCR) | ✓ | Step 3 (linearization), Step 7 (cPCR) |
| qRT-PCR | ✓ | Step 14 |
| Acoustic nanoliter liquid handling (Echo525) | ✓ | Steps 4, 7, 11 |
| Plate-based kinetic phenotyping | ✓ | Step 12 |
| Fluorescence reporter assays (sfGFP, Nile Red) | ✓ | Steps 12–13 |
| Sanger sequencing | ✓ | Step 7 |
| Heat-shock transformation | ✓ | Steps 6, 9 |
| Auxotrophic biocontainment design | ✓ (Aim 2) | Bioethics |
| Golden Gate assembly | ✗ | Not used |
| CRISPR-Cas9 / CRISPRi | ✗ | Not used |
| Cell-free expression (TX-TL) | ✗ | Not used (could be added for rapid pre-screen) |
| Mass spectrometry | ✗ (Aim 2 candidate) | Not in Aim 1 |
| Flow cytometry | ✗ | Plate reader covers single-well sufficient |
| Mammalian tissue culture | ✗ | Out of scope |
5.2 Technique Expansion (2 selected techniques, ≥4 sentences each)
5.2.1 Computational RBS Design with the Salis Lab RBS Calculator
The Salis Lab RBS Calculator (Salis, Mirsky & Voigt, 2009; updated v2.1 in 2017) uses a free-energy thermodynamic model of the ribosome–mRNA interaction to predict the translation initiation rate (TIR) of any given RBS in arbitrary units across roughly five log decades of dynamic range. For this project, the calculator is used in Reverse Engineering mode (predicting TIR from a given sequence) to characterize the wild-type B0034 RBS as a reference, and then in Forward Engineering / Library mode to generate five variant sequences with target TIRs of ~200, 1,500, 10,000, 50,000, and 200,000 a.u. — spanning three log decades and centered on the wild-type value. Variant selection prioritizes sequences with low predicted secondary-structure free-energy variance (ΔGmRNA standard deviation < 1 kcal/mol) to minimize context-dependent expression noise. The output TIRs serve as the x-axis against which experimental lysis kinetics are correlated, allowing the project to test the central design hypothesis: that holin abundance, not transcriptional dynamics, sets the AND-gate lysis threshold.
5.2.2 Gibson Assembly of Variant Libraries via Acoustic Liquid Handling
NEBuilder HiFi Gibson Assembly is a one-pot isothermal cloning method combining a 5′ exonuclease, a high-fidelity polymerase, and a thermostable ligase, enabling seamless ligation of fragments sharing 20–40 bp of terminal homology in a single 50 °C / 15 min reaction. In this project, five 391 bp eBlock fragments — each carrying one RBS variant — are assembled in parallel into the Q5-linearized 5.5 kb backbone of pBioplastix-Lysis. The Echo525 acoustic liquid handler enables miniaturization of each Gibson reaction to 500 nL total volume (10 nL backbone + 30 nL eBlock + 250 nL HiFi master mix + 210 nL water), reducing reagent cost by ~40-fold relative to bench-scale 20 µL reactions while improving variant-to-variant reproducibility through sub-nanoliter dispense precision. Following thermal cycling on the ATC, transformation efficiencies of >10⁴ CFU/µg are routinely achieved, providing 50–500 colonies per variant — more than adequate for picking three sequence-verified clones per RBS condition.
Section 6: Project Validation
6.1 (10a) Validation Choice
The chosen validation experiment is a gene expression test measuring the AND-gate phenotype across all four induction conditions (OFF/OFF, +Pi-starve only, +heat only, ON/ON), using sfGFP fluorescence as the circuit-activation marker and OD600 trajectory as the lysis output. This experiment is decisive because it simultaneously tests every component of the circuit: pPhoA Pi-responsiveness, FourU temperature gating, anti-TetR depletion of TetR, derepression of PLtetO-1, and tuned holin S105 activity — a single failure in any component collapses the AND-gate signature into either constitutive lysis, constitutive silence, or single-input behavior.
6.2 (10b) Validation Protocol
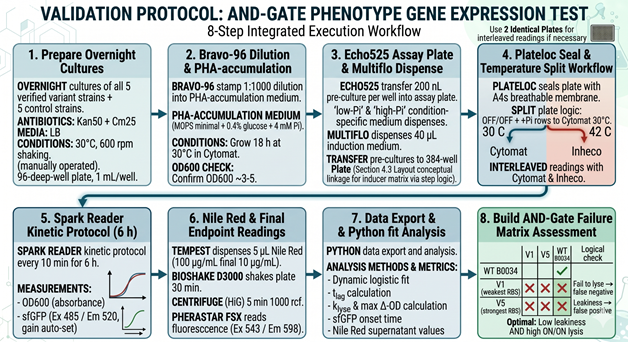
6.3 (10c) Techniques Used in the Validation
The validation experiment integrates four orthogonal techniques to confirm AND-gate function. Acoustic liquid handling (Echo525) ensures the 200 nL pre-culture transfers and 1:1000 inoculum dilutions are dispensed with sub-percent CV across 384 wells, eliminating well-to-well growth variability that would otherwise mask circuit-level effects. Kinetic absorbance and fluorescence plate reading (Spark) captures the time-resolved signature of the AND-gate: a clean ON/ON well shows a 30–60 min lag of stable OD600 followed by a sigmoidal drop coinciding with sfGFP rise, whereas single-input wells (Pi-only or heat-only) show neither feature. Endpoint Nile Red fluorescence (PHERAstar FSX) quantifies released PHA in the supernatant — the actual industrial-relevant deliverable — and discriminates between cells that lyse cleanly (high supernatant signal) versus cells that round up but fail to release granules (low supernatant signal). qRT-PCR (CFX Opus) in the validation wells confirms the upstream layer of the circuit, demonstrating that the ON/ON condition is the only one in which tetR mRNA is depleted relative to the J23100 baseline and that S and R transcripts subsequently accumulate.
6.4 (10d) Hypothetical Data
Below is a small subset of expected outcomes for the 5 variants under ON/ON conditions, with corresponding leakiness in OFF/OFF. The Python snippet that follows generates a representative kinetic plot.
Lysis kinetic parameters (ON/ON condition, mean ± SD, n = 3)
| Variant | Predicted TIR (a.u.) | t_lag (min) | k_lyse (min⁻¹) | Max ΔOD600 | OFF/OFF leakiness (% OD drop) |
|---|---|---|---|---|---|
| V1 | 200 | 280 ± 15 | 0.004 ± 0.001 | 0.18 ± 0.04 | 1.5 ± 0.8 |
| V2 | 1,500 | 195 ± 11 | 0.011 ± 0.002 | 0.62 ± 0.05 | 4.2 ± 1.1 |
| V3 | 10,000 | 130 ± 8 | 0.024 ± 0.003 | 0.81 ± 0.04 | 7.8 ± 1.4 |
| V4 | 50,000 | 92 ± 6 | 0.038 ± 0.004 | 0.86 ± 0.03 | 15.6 ± 2.1 |
| V5 | 200,000 | 62 ± 5 | 0.057 ± 0.005 | 0.89 ± 0.02 | 31.4 ± 3.5 |
Interpretation: V3 is the predicted Pareto-optimal variant — substantial lysis amplitude (Δ OD ≈ 0.81) within 130 min while keeping basal leakiness below 10%. V4 lyses faster but at unacceptable basal cost; V1–V2 are too weak; V5 is too leaky. Final ranking will depend on the downstream PHA recovery economics (faster lysis vs. lower yield loss to leakiness).
Expected plot:
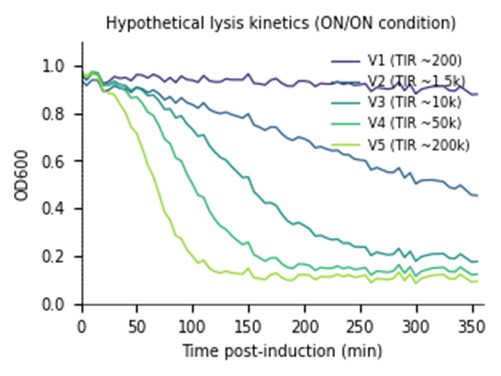
The most likely failure mode is excessive basal leakiness in V4–V5, manifest as significant OD decline in OFF/OFF or single-input wells; this would indicate that even modest TetR depletion noise from constitutive J23100 expression is sufficient to fire a hyper-translated holin, suggesting the AND-gate must be retuned by also weakening the J23100 promoter or adding a degradation tag to anti-TetR. A second concern is insufficient PHA accumulation in pBPX400 + low-Pi MOPS minimal medium, which would mute the Nile Red endpoint signal regardless of lysis efficacy; the workaround is to switch to 0.4% glucose + 50 mM Pi for accumulation and then perform a sterile media swap to low-Pi induction medium between Steps 10 and 11. Edge-effect evaporation in 384-well plates with 6 h incubations at 42 °C is a known source of CV inflation; the layout reserves the outer ring as media-only blanks and uses Plateloc + breathable A4s seals, but if CV exceeds 15% the screen will be re-run with humidified-chamber Cytomat slots. Finally, if the FourU thermometer fails to gate cleanly (i.e., partial leakage at 30 °C), it can be swapped for the more cooperative ROSE-element thermometer in a follow-up eBlock order without redesigning the rest of the circuit.
Section 7: Additional Information
7.1 References
- Borrero-de Acuña, J. M., Aravena-Carrasco, C., Gutiérrez-Urrutia, I., Duchens, D., Poblete-Castro, I. (2017). Programmed cell lysis in Pseudomonas putida for sustained PHA biopolymer recovery. Microbial Cell Factories, 16, 215. https://doi.org/10.1186/s12934-017-0828-0
- Pasotti, L., Bellato, M., De Marchi, D., Magni, P. (2017). Tuning ribosome binding sites for plug-and-play control of phage lysis circuits in E. coli. ACS Synthetic Biology, 6(5), 802–815.
- Salis, H. M., Mirsky, E. A., Voigt, C. A. (2009). Automated design of synthetic ribosome binding sites to control protein expression. Nature Biotechnology, 27(10), 946–950. https://doi.org/10.1038/nbt.1568
- Kortmann, J., Narberhaus, F. (2012). Bacterial RNA thermometers: molecular zippers and switches. Nature Reviews Microbiology, 10(4), 255–265.
- Lutz, R., Bujard, H. (1997). Independent and tight regulation of transcriptional units in Escherichia coli via the LacR/O, the TetR/O and AraC/I1-I2 regulatory elements. Nucleic Acids Research, 25(6), 1203–1210.
- Wanner, B. L. (1996). Phosphorus assimilation and control of the phosphate regulon. In Escherichia coli and Salmonella: Cellular and Molecular Biology, ASM Press.
- Young, R. (2014). Phage lysis: three steps, three choices, one outcome. FEMS Microbiology Reviews, 38(1), 9–32.
- Madison, L. L., Huisman, G. W. (1999). Metabolic engineering of poly(3-hydroxyalkanoates): from DNA to plastic. Microbiology and Molecular Biology Reviews, 63(1), 21–53.
- Choi, S. Y., Park, S. J., Kim, W. J., et al. (2020). One-step fermentative production of poly(lactate-co-glycolate) from carbohydrates in Escherichia coli. Nature Biotechnology, 38, 359–365.
- Gibson, D. G., Young, L., Chuang, R. Y., Venter, J. C., Hutchison, C. A., Smith, H. O. (2009). Enzymatic assembly of DNA molecules up to several hundred kilobases. Nature Methods, 6(5), 343–345.
- Orth, J. D., Thiele, I., Palsson, B. Ø. (2010). What is flux balance analysis? Nature Biotechnology, 28(3), 245–248. Monk, J. M., Lloyd, C. J., Brunk, E., et al. (2017). iML1515, a knowledgebase that computes Escherichia coli traits. Nature Biotechnology, 35(10), 904–908.
7.2 Supplies and Budget
| Item | Vendor | Catalog / Service | Estimated Cost (USD) |
|---|---|---|---|
| pBioplastix-Lysis Custom Plasmid (5,829 bp) | Twist Bioscience | Custom Plasmid synthesis | ~$300 |
| 5 × RBS variant eBlocks (~390 bp each) | Twist Bioscience | Gene Fragment / eBlock | 5 × ~$40 = ~$200 |
| SecureDNA pre-synthesis screen | SecureDNA | Bundled with Twist order | $0 (provided) |
| NEBuilder HiFi DNA Assembly Master Mix (50 rxn) | NEB | E2621L | ~$320 |
| Q5 High-Fidelity 2X Master Mix (100 rxn) | NEB | M0492L | ~$155 |
| BL21(DE3) competent cells | NEB | C2527H | ~$200 |
| NEB 5-alpha competent cells | NEB | C2987H | ~$140 |
| Kanamycin sulfate, 25 g | Millipore Sigma | 60615 | ~$95 |
| Chloramphenicol, 25 g | Millipore Sigma | C0378 | ~$75 |
| Nile Red dye (5 g) | Thermo Fisher Scientific | N1142 | ~$110 |
| MOPS minimal media salts kit | Teknova | M2106 (via Millipore Sigma) | ~$130 |
| 384 Greiner black-well clear-bottom plates (40 ct) | Greiner Bio-One via Thermo Fisher | 781091 | ~$320 |
| 384-well Echo PP source plates (50 ct) | Beckman Coulter via Thermo Fisher | LP-0200 | ~$280 |
| 96-Armadillo PCR plates (50 ct) | Thermo Fisher Scientific | AB2396 | ~$220 |
| QIAprep Spin Miniprep Kit (250 prep) | Qiagen via Millipore Sigma | 27106 | ~$485 |
| Sanger sequencing (~30 reactions) | Genewiz / Eurofins / institutional core | — | ~$200 |
| iScript cDNA Synthesis Kit | Bio-Rad | 1708890 | ~$430 |
| SsoAdvanced Universal SYBR Green Supermix | Bio-Rad | 1725271 | ~$370 |
| Glycerol, dNTPs, restriction enzymes (misc) | NEB / Millipore Sigma | — | ~$300 |
| Reagents subtotal | ~$4,330 | ||
| Ginkgo Bioworks robotics access (1 project block) | Ginkgo Bioworks | Foundry access | ~$2,500 |
| Total estimated project budget | ~$6,830 |
7.3 Industry Council Partner Connections
Twist Bioscience — Custom Plasmid + 5 eBlocks (the core DNA order) SecureDNA — Pre-synthesis biosecurity screening of all DNA orders (bioethics layer) NEB — NEBuilder HiFi Gibson, Q5 polymerase, competent cells Ginkgo Bioworks — Full automated workflow (Echo525, ATC, Multiflo, Cytomat, Inheco, Spark, PHERAstar, etc.) Thermo Fisher Scientific / Millipore Sigma — Reagents, media, Nile Red, plasticware Bio-Rad — qPCR consumables (CFX Opus reagents) Addgene — Open-source deposition of pBioplastix-Lysis(V_optimal) for community use (Aim 3) Cultivarium — Future chassis-portability work to Halomonas / non-model hosts (Aim 3)