Week 11 HW: Bioproduction & Cloud Labs
Week 11 : Bioproduction & Cloud Labs
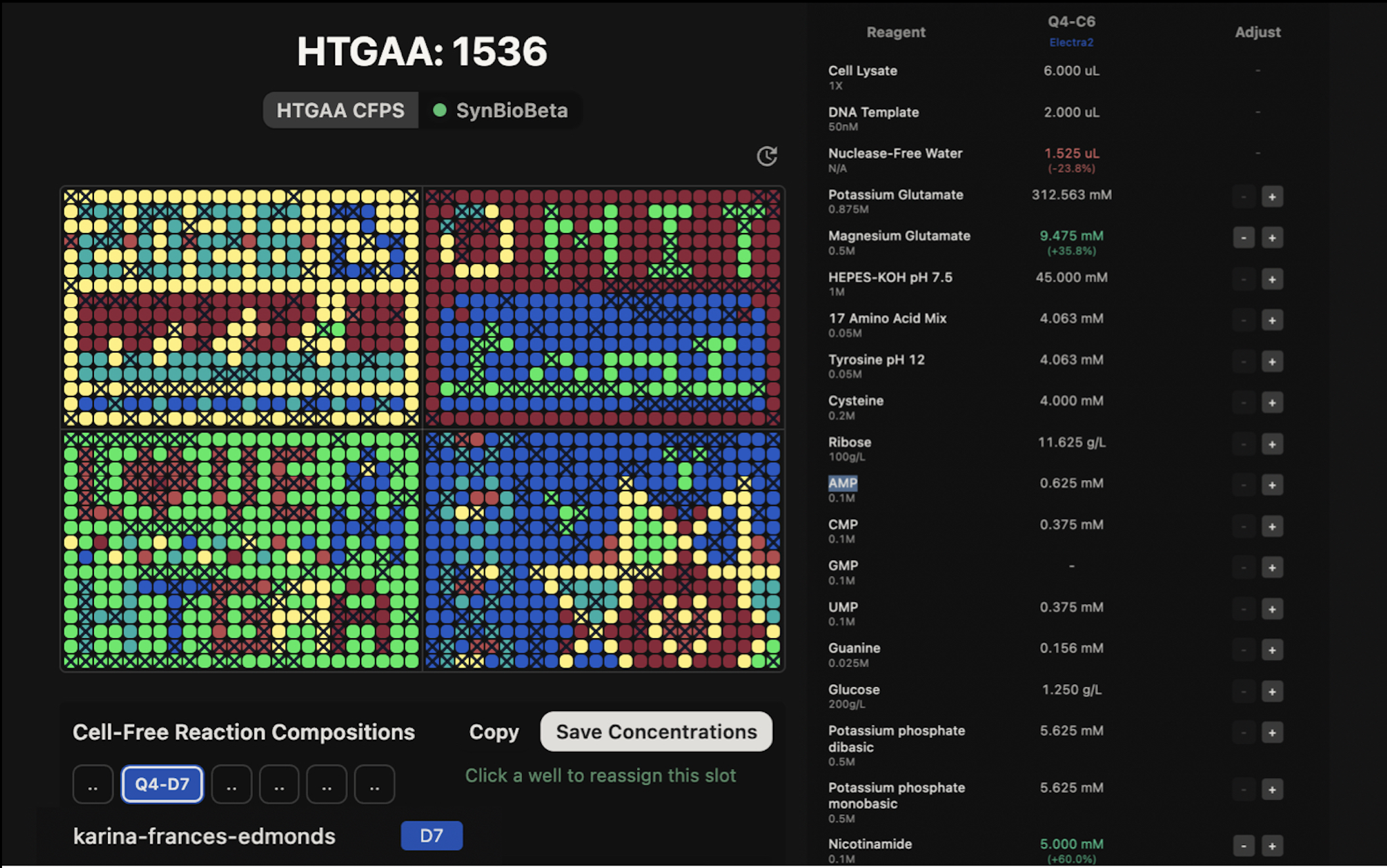
Part A: The 1,536 Pixel Artwork Canvas | Collective Artwork
Code Formula [ { “quadrant”: “Q4”, “well_label”: “D6”, “supplements”: [ { “id”: “nuclease_free_water”, “supplemental_volume_nl”: 1250 }, { “id”: “glucose”, “supplemental_volume_nl”: 125 }, { “id”: “nicotinamide”, “supplemental_volume_nl”: 625 } ] } ]
Findings
I found the Collective Cloud Artwork really interesting, I liked the idea of collaborating not just on art but on biology simultaneously, like each person contributing one ingredient to a shared recipe, or a single cell within a larger organism. There are so many cool potential applications for this kind of collective experiment. I personally appreciated the concept of each person adding only one well, since it reduces individual control and makes the outcome more spontaneous and truly collaborative. The fact that every student’s composition is slightly different also means the artwork doubles as a large-scale reagent optimisation experiment, when the fluorescence data comes back, correlating pixel brightness with each person’s reagent choices could reveal genuinely useful design rules for cell-free systems.
I contributed to the cell-free fluorescent protein artwork by composing a custom reagent supplement for well Q4-C6, assigned to the Electra2 far-red fluorescent protein. My composition doubled both Glucose (1.25 → 2.5 g/L) and Nicotinamide (3.125 → 6.25 mM) relative to the baseline 20-hour NMP-Ribose master mix, targeting sustained NAD⁺ regeneration through two independent metabolic routes over the full 36-hour incubation.
Future Improvements
Giving students more than 2 µL of supplement volume, or allowing adjustments to more reagents, would widen the hypothesis space and make the resulting dataset more statistically powerful. Perhaps there could also be an AI assistant that could give feedback in real time based on the reagent trials. Perhaps the student can have freedom with the flourescent choice, giving more of an artistic exploration.
Roles
E. coli Lysate BL21(DE3) Star Provides the complete cellular machinery for gene expression: ribosomes, translation factors, tRNA synthetases, chaperones, and T7 RNA polymerase for transcription of the DNA template.
Potassium Glutamate - 312.6 mM Maintains physiological ionic strength and stabilizes ribosome structure; acts as a compatible solute mimicking the intracellular environment of E. coli.
HEPES-KOH pH 7.5 - 45 mM Buffers the reaction at pH 7.5, the optimal pH for ribosome activity and most translation factors.
Magnesium Glutamate - 7.0 mM Mg²⁺ is essential for ribosome assembly, peptidyl-transferase activity, and stabilizing RNA secondary structures throughout transcription and translation.
Potassium phosphate dibasic + monobasic - 5.6 mM each Provides inorganic phosphate as a secondary pH buffer and phosphate donor for NMP-to-NTP phosphorylation reactions.
Ribose - 11.625 g/L (~77.4 mM) The pentose sugar backbone for nucleotide synthesis; lysate kinases convert ribose into phosphorylated intermediates that regenerate NTPs from NMPs sustainably over long incubations.
Glucose - 1.25 g/L (baseline) → 2.5 g/L (our supplement) Feeds glycolysis to actively regenerate ATP and NAD⁺ via GAPDH, providing a second independent energy source alongside the NMP-ribose recycling system.
AMP - 0.625 mM | CMP · 0.375 mM | UMP · 0.375 mM Nucleoside monophosphates that are phosphorylated by lysate kinases to their active triphosphate forms for RNA synthesis and energy metabolism.
GMP - 0 µM Absent because GTP is instead generated from free guanine via the purine salvage pathway — a cheaper and more stable alternative.
Guanine - 0.156 mM Salvaged by HGPRT in the lysate and converted to GMP, then phosphorylated to GTP, making it available for transcription without adding GMP directly.
17 Amino Acid Mix - 4.063 mM Supplies all standard amino acids except tyrosine and cysteine, providing the building blocks for polypeptide synthesis.
Tyrosine pH 12 - 4.063 mM Added separately at high pH because tyrosine is nearly insoluble at neutral pH; dissolved at pH 12 to maintain concentration without precipitating.
Cysteine - 4.000 mM Added separately because cysteine oxidizes rapidly and can form unwanted disulfide crosslinks if premixed; also critical for proper beta-barrel folding in many fluorescent proteins.
Nicotinamide - 3.125 mM (baseline) → 6.25 mM (our supplement) Inhibits NADase activity in the lysate to preserve NAD⁺ levels needed for energy metabolism; we double this to maximize NAD⁺ protection over the full 36-hour incubation.
Nuclease-Free Water (backfill) Brings the master mix to final reaction volume; RNase/DNase-free to prevent degradation of the DNA template and mRNA transcripts.
Bonus
Although GMP is absent, the lysate contains the purine salvage enzyme HGPRT, which attaches free guanine to PRPP to form GMP on demand. GMP is then sequentially phosphorylated to GTP by nucleoside kinases already present in the lysate, making free guanine a cheaper and more stable way to supply GTP for transcription without adding it directly.
Part B : 1H PEP-NTP VS 20H NMP-Ribose Differences
The 1-hour system gives the reaction everything needed for the reaction to occur. All four building blocks for RNA (NTPs) plus fast energy sources, thus proteins are made quickly however the reaction burns out within an hour. The 20-hour system instead provides simpler ingredients that the lysate’s own enzymes slowly convert into NTPs as needed, like a slow-release fuel, keeping the reaction running sustainably for much longer. It’s also a cleaner, simpler recipe, fewer additives, and only includes the essentials.
Protein: Electra2 (far-red, slow chromophore maturation)
Property targeted: Sustained translational activity and NAD⁺ homeostasis over 36 hours
Hypothesis: The hypothesis is that doubling both Glucose (1.25 → 2.5 g/L) and Nicotinamide (3.125 → 6.25 mM) will maximise Electra2 fluorescence over a 36-hour cell-free incubation by sustaining NAD⁺ availability through two independent metabolic routes. Nicotinamide inhibits NADase in the lysate, slowing passive NAD⁺ degradation, while elevated Glucose feeds glycolysis to actively regenerate NAD⁺ via GAPDH, together maintaining the redox balance required for continuous NTP recycling from NMPs. Because Electra2 is a slow-maturing far-red protein whose chromophore cyclisation occurs late in the incubation, sustained translational activity over the full 36 hours is more critical than for fast-maturing proteins like sfGFP, making long-term energy regeneration the primary bottleneck to address. This dual-pathway strategy is the most distinctive approach to solving it.
Part D:
Building a Cloud Lab

I was not successful in the optional assignment, but I genuinely enjoyed exploring the models across their different view frames and layouts. What I find fascinating is how biology is increasingly becoming a designable, digital medium, and as someone who works at the intersection of design, biology and technology, that feels deeply relevant to my own practice. The more we digitise and standardise these systems, the more I believe knowledge can be shared universally, making science less siloed and more collectively accessible.