Week 11 HW: Bioproduction and Cloud Labs

Part A: The 1,536 Pixel Artwork Canvas | Collective Artwork
My Contribution
My involvement in the community bioart project started right at the beginning. I found myself in a “digital battle” with other users over whether the top row on the left plate would be blue or yellow - I was on Team Yellow! No particular reason, just a gut feeling:) After a few minutes, others joined me:
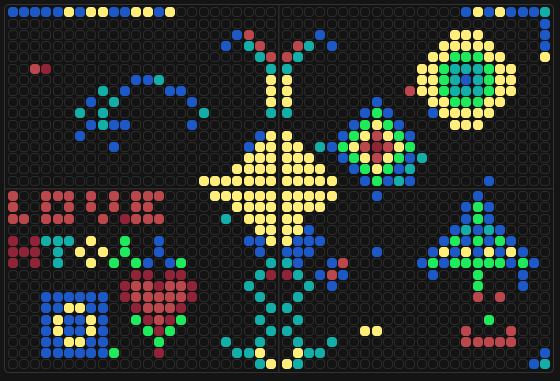
It did indeed end up yellow in the final design!
What I liked
I really enjoyed the global aspect of the project - the fact that hundreds of people from all around the world contributed to a single piece of art is remarkable. I also loved the “evolutionary” dimension of it: being able to trace how the design changed and developed over time gave it a unique, living quality.
My favourite snapshot from the evolution:
What could be improved
For next year, I’d suggest two things:
- More colors - a broader palette would give participants more creative flexibility and make the process even more fun.
- More surface area - a larger canvas would allow for higher-resolution details and give more people the space to leave their mark on the final piece.
Overall, it was a wonderful idea and a genuinely fun experience to be part of!
Part B: Cell-Free Protein Synthesis | Cell-Free Reagents
1. Cell-Free Protein Synthesis Reaction Components
E. coli Lysate
| Component | Role |
|---|---|
| BL21 (DE3) Star Lysate (includes T7 RNA Polymerase) | Provides the core cellular machinery needed for transcription and translation, including ribosomes, enzymes, and co-factors. The T7 RNA Polymerase specifically transcribes DNA into mRNA using the T7 promoter system. |
Salts / Buffer
| Component | Role |
|---|---|
| Potassium Glutamate | Maintains ionic strength and mimics the intracellular environment, supporting enzyme activity and ribosome stability. |
| HEPES-KOH pH 7.5 | A buffering agent that maintains a stable, physiological pH (7.5), which is optimal for transcription and translation enzymes. |
| Magnesium Glutamate | Provides Mg²⁺ ions, which are essential cofactors for ribosomes, RNA polymerase, and many enzymes involved in protein synthesis. |
| Potassium Phosphate Monobasic | Works together with the dibasic form to maintain pH and provide phosphate ions, which are important for energy metabolism and nucleotide synthesis. |
| Potassium Phosphate Dibasic | Works together with the monobasic form to maintain pH and provide phosphate ions, which are important for energy metabolism and nucleotide synthesis. |
Energy / Nucleotide System
| Component | Role |
|---|---|
| Ribose | A simple sugar that serves as a carbon and energy source; can be metabolized to regenerate nucleotides and sustain the reaction over longer incubation periods. |
| Glucose | Another energy source that feeds into central metabolic pathways to regenerate ATP and other nucleotides needed for sustained transcription and translation. |
| AMP | A nucleotide monophosphate precursor that is phosphorylated to ATP, the primary energy currency that drives protein synthesis. |
| CMP | Precursor to CTP, which is required for RNA synthesis during transcription. |
| GMP | Precursor to GTP, which is required for both transcription and translation (e.g., ribosome translocation). |
| UMP | Precursor to UTP, which is required for RNA synthesis during transcription. |
| Guanine | A nucleobase that can be salvaged and converted into GMP/GTP, supplementing the guanosine nucleotide pool for transcription and translation. |
Translation Mix (Amino Acids)
| Component | Role |
|---|---|
| 17 Amino Acid Mix | Provides the bulk of the amino acid building blocks needed for the ribosome to synthesize polypeptide chains. |
| Tyrosine | Supplied separately because of its low solubility at neutral pH; it is an essential amino acid for building complete proteins. |
| Cysteine | Supplied separately due to its reactivity; it is essential for forming disulfide bonds and maintaining the structural integrity of many proteins. |
Additives
| Component | Role |
|---|---|
| Nicotinamide | A precursor to NAD⁺, which is a critical cofactor in metabolic redox reactions that regenerate energy to sustain the cell-free reaction. |
Backfill
| Component | Role |
|---|---|
| Nuclease-Free Water | Used to bring the reaction to its final volume without introducing RNases (ribonucleases) and DNases (deoxyribonucleases), enzymes that degrade RNA and DNA respectively, that could degrade the nucleic acids in the reaction. |
2. 1-Hour PEP/NTP vs. 20-Hour NMP-Ribose Master Mix: Key Differences
The most fundamental difference between the two master mixes is their energy and nucleotide system: the 1-hour PEP/NTP mix supplies energy directly via pre-made NTPs (ATP, GTP, CTP, UTP) and PEP-Mono (phosphoenolpyruvate monopotassium salt; regenerates NTPs by donating phosphate groups to NDPs produced during transcription) as an immediate energy source, while the 20-hour NMP-Ribose mix supplies energy indirectly via NMPs and simple sugars (ribose and glucose) that the lysate enzymes must first convert into usable NTPs. This makes the PEP/NTP system fast but short-lived, while the NMP-Ribose system is slower to get going but more sustainable over longer incubation periods. The 1-hour mix also contains more additives (Spermidine, DMSO, cAMP, NAD, Folinic Acid) to boost immediate transcription/translation activity, whereas the 20-hour mix keeps additives minimal (only Nicotinamide) since the reaction has more time to proceed on its own.
3. Bonus question
Although GMP is listed as 0.00 uM in the 20-hour mix, Guanine (the nucleobase) is still provided and can be converted into GMP through a process called nucleotide salvage, where enzymes in the lysate attach a ribose-phosphate group to the free Guanine base. The resulting GMP is then phosphorylated up to GTP, which can be used for transcription. This is a more cost-effective way to supply guanosine nucleotides compared to adding pre-made GMP directly.
Part C: Planning the Global Experiment | Cell-Free Master Mix Design
A. Biophysical and Functional Properties of the 6 Fluorescent Proteins
1. sfGFP (Superfolder GFP)
sfGFP is engineered to fold extremely rapidly and robustly, even when fused to poorly folding proteins, making it one of the most reliable reporters in cell-free systems. Like all GFP-based proteins, its chromophore maturation requires molecular oxygen, meaning fluorescence will be delayed or absent in low-oxygen conditions.
2. mRFP1 (Monomeric Red Fluorescent Protein 1)
mRFP1 is a true monomer, which is important in cell-free systems because oligomerizing proteins can cause aggregation artifacts. However, it has relatively low brightness and photostability compared to newer red fluorescent proteins, which can limit signal readout in cell-free reactions where protein yields may already be low.
3. mKO2 (Monomeric Kusabira Orange 2)
mKO2 is an orange fluorescent protein with moderate acid sensitivity, meaning its fluorescence can be partially quenched if the pH of the cell-free reaction drifts below neutral. It also has a relatively slow and complex maturation process (requiring both a folding and a separate maturation step), which can delay fluorescence readout in cell-free systems.
4. mTurquoise2
mTurquoise2 is a rapidly maturing cyan fluorescent protein with very low acid sensitivity, making it a stable and reliable reporter across different reaction conditions. Its fast maturation is particularly advantageous in cell-free systems, where getting a quick fluorescence readout is important.
5. mScarlet-I
mScarlet-I is a synthetically derived, rapidly maturing monomeric red fluorescent protein with high brightness, making it one of the best-performing red reporters in cell-free systems. Like other GFP-family proteins, it requires oxygen for chromophore maturation, so adequate oxygenation of the reaction is important for good signal.
6. Electra2
Electra2 is a blue fluorescent protein (excitation ~405 nm, emission ~456 nm) derived from Entacmaea quadricolor, engineered for high intracellular brightness. Its blue emission provides good spectral separation from green and red fluorescent proteins, making it useful in multicolor cell-free experiments — though blue fluorescent proteins generally have lower brightness than their green and red counterparts, which can affect detection sensitivity.
B. Cell-Free Optimization Experiment: mRFP1 vs mScarlet-I over 36 Hours
Hypothesis
mRFP1’s main limitations are slow maturation and low brightness. I hypothesize that supplementing the 36-hour master mix with additional Cysteine (critical for mRFP1’s disulfide bonds and folding) and GMP (to boost GTP availability for sustained transcription) will improve folding efficiency and protein yield, thereby maximizing fluorescence over 36 hours. I will use mScarlet-I as a positive control reference since it is a fast-maturing red protein, to see whether my optimization can close the brightness gap.
Experimental Design
mRFP1 Wells (4 wells — varying Cysteine + GMP)
| Well | Protein | GMP added | Cysteine added | Notes |
|---|---|---|---|---|
| A1 | mRFP1 | 0 mM | 0 mM | Baseline control |
| A2 | mRFP1 | 0.250 mM | 0 mM | GMP only |
| A3 | mRFP1 | 0 mM | 0.500 mM | Cysteine only |
| A4 | mRFP1 | 0.250 mM | 0.500 mM | Combined |
mScarlet-I Wells (4 wells — same design, for comparison)
| Well | Protein | GMP added | Cysteine added | Notes |
|---|---|---|---|---|
| B1 | mScarlet-I | 0 mM | 0 mM | Baseline control |
| B2 | mScarlet-I | 0.250 mM | 0 mM | GMP only |
| B3 | mScarlet-I | 0 mM | 0.500 mM | Cysteine only |
| B4 | mScarlet-I | 0.250 mM | 0.500 mM | Combined |
All wells contain: 6 µL Lysate + 10 µL 2X Master Mix + 2 µL DNA template + 0–2 µL custom supplements (backfilled with nuclease-free water to 2 µL)
I input these concentrations on the website
Reagent Rationale
GMP → GTP: The baseline 36-hour mix has GMP = 0, relying entirely on Guanine salvage. Adding a small GMP boost gives the system more immediate GTP for transcription, which should increase mRNA production early in the reaction and improve total protein yield.
Cysteine: mRFP1 underwent 33 mutations during its engineering and contains cysteines important for proper folding. The baseline Cysteine concentration is 4.00 mM — adding 0.25 mM is a modest, safe increase that could improve folding efficiency without disrupting the reaction balance.
A1 and B1 as controls: Essential baselines to compare against — if A4 is brighter than A1, the supplementation worked.
Expected Outcome
I expect fluorescence in the mRFP1 wells to follow the order A4 > A3 > A2 > A1, with the combined GMP + Cysteine well giving the best result. The mScarlet-I wells (B1–B4) are expected to be brighter than their mRFP1 counterparts overall, given mScarlet-I’s faster maturation and higher intrinsic brightness. However, the supplements may also boost mScarlet-I fluorescence - comparing B1 vs B4 will reveal whether these additions benefit fast-maturing proteins as much as slow-maturing ones like mRFP1.