PHOTOSYNTHETIC IMAGE GENERATION: Co-Cultured Autotrophic-Heterotrophic Color Controls for Bacteria Based Imagery
HTGAA TA’s: Danny Chen, Inger Le Gué, Juan Diego Unda, Digby Usher
SECTION 1: ABSTRACT:
Currently, biological art and engineered living material systems are often based on heterotrophic microorganisms such as E. coli, which require continuous external nutrient supplementation to function. In contrast this project looks at cyanobacteria as a potential solution to nutrient supply since they are capable of using light and CO₂ to naturally generate organic carbon through photosynthesis. However, cyanobacteria is less suitable for engineering visual outputs. The goal of this project is to combine cyanobacteria and E. coli into a synthetic co-culture, where cyanobacteria generates sucrose to support engineered E. coli for the production of chromo-proteins in a continuously recirculating visual media system.
SECTION 2: PROJECT AIMS
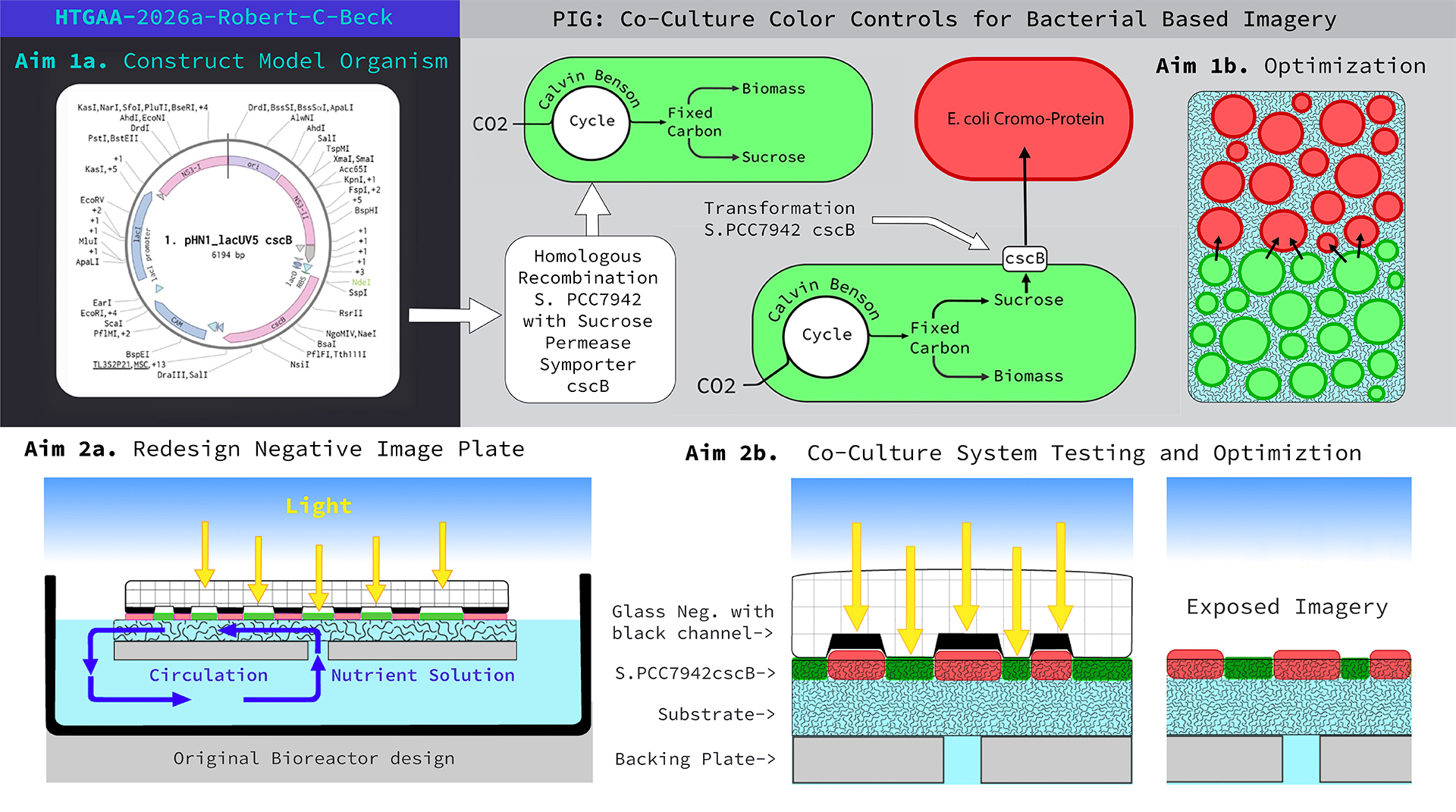
Aim I: Design and construct plasmids for homologous recombination of cyanobacterial strain, Synechococcus elongatus PCC 7942 with the sucrose permease cscB gene for secretion of sucrose.
Design functional cscB plasmids and submit order to Twist.
With arrival of plasmids, complete first transformation of S. 7942, and transfer microorganism to cold storage.
Redesign negative image plate for co-culture bioreactor.
Aim II: The co-culture system will require optimization of environment and media before attempting the co-culture.
First subaim- will test culture of S. 7942 cscB+ under control of salt stress and IPTG for sucrose production. Control variables will include, but are not limited to, the following:
- Nutrient concentrations of BG11 media.
- NaCl induced salt stress vs. KCl.
- Growth under LED and fluorescent lighting.
Second subaim- builds on results from the optimized media tests to construct the co-culture system with E. coli chromo-protein.
Third subaim- fabricates the bio-reactor negative image plates from Aim 1.
Aim III: Multi color expressions in the co-culture system, alternative salt free cyanobacteria systems, and mobile lab construction.
First subaim- tests multiple E. coli chromo-proteins within the co-culture system to generate multi color expressions.Design and fabricate mobile lab system for transportation of work to exhibition venues.
Second subaim- Design and fabricate mobile lab system for transportation of work to exhibition venues.
Third subaim- exhibits the project outside of the lab in alternative setting.
SECTION 3: BACKGROUND
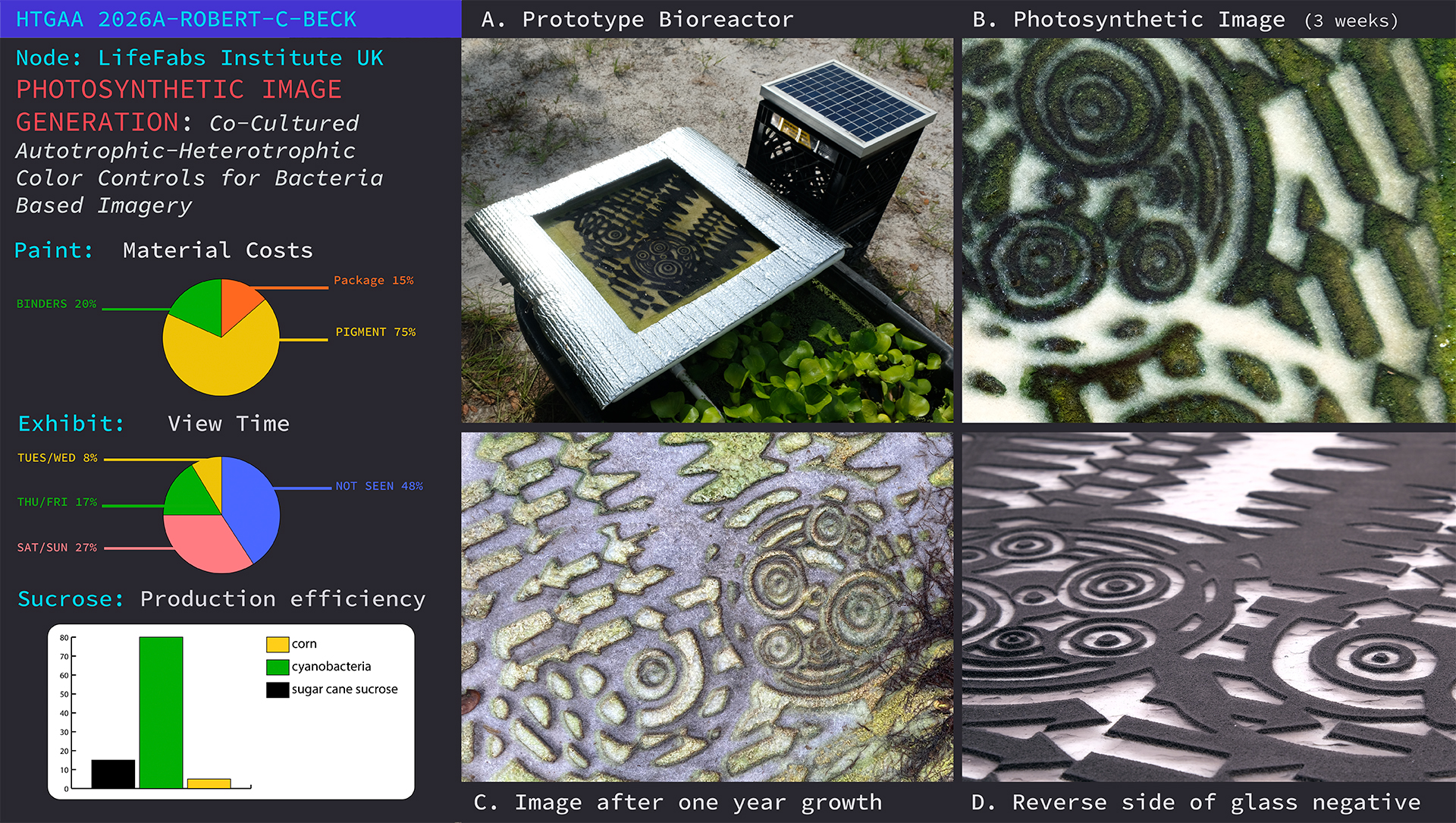
Based on research of co-culturing Cyanobacteria Synechococcus elongateus 7942 cscB (S. cscB 7942) with Escherichia coli for bio-chemical synthesis [1,2], this project will explore the adaptation of the co-culturing process to produce visual color modifications within an image by generating muti-colored bacteria based pixels in a continuous reaction. The origin of this theoretical approach to image cultivation began through a series of visual experiments in collaboration with my partner, Sarah Max Beck, entitled studioHydrostatic. The project was set on top of a Brooklyn rooftop where we were experimenting with aqauponic technology to create a recirculating, human based, organic food production system. Two large flood tables grew a variety of edible plants and herbs in nitrogen rich water. This is where I first discovered a blue-green microorganism growing in a polka-dot like pattern on a sheet of bubble-foil insulation within the system. Recognition of the pattern lead to the developent of a glass plate negative (figure 2) for growing wild caught cyanobacteria in a contolled visual experiment.
Field tests were conducted in 2020 -2021, which proved the viability of the concept. It was during this initial trial when the first full size plate, WTF, was tested in a prototype bioreactor producing a graphic image (figure 1). As the cyanobacteria growth continued to progress in the WTF image, the density of bacteria progressively darkened the image. It then started to build on itself creating a slightly elevated surface of growth; it appeared to be different from the cyanobacteria. As it progressed the biomass reached a thickness of aproximately 2-3 mm after one year of growth. Eventually, it took over the image, and significantly suppressed the cyanobacteria’s chlorophyll pigmentation (figure 3). While I can only speculate, I believe the image was overtaken by some form of E. coli judging from the color and texture of the biomass.
My speculation is based on lab grown E. coli cultures observed during HTGAA labs with Lifefabs Institute, which indicated there’s a natural relationship between these organisms. Searching for evidence to support the connection, the paper entitled, Cross-feeding between cyanobacterium Synechococcus and Escherichia coli in an artificial autotrophic-heterotrphic cocuture system revealed by integrated omic analysis, demonstrated that cyanobacteria could be engineered to export sucrose and provide energy for E. coli [1].
The possibility of creating a symbiotic relationship between two different organisms to enhance the graphic functionality of PIG is the driving motivation behind this project. With the discovery of the co-culture technique came the task of understanding the mechanisms behind creating a similar relationship, where cross-feeding would result in a multi-color image of chromo-proteins supported by cyanobacteria’s ability to supply carbon to E. coli for the production of chromo-protein. In doing so, this project will reveal a different relationship between these organisms, and advance the field of biological art with a new system for growing visual media, which exceeds the time limitations of gel art, by co-culturing organisms under the control of a photonegative plate engineered specifically for this process.
PAPER #1:Cross-feeding between cyanobacterium Synechococcus and Escherichia coli in an artificial autotrophic-heterotrophic coculture system revealed by integrated omic analysis
In aquatic environments, the ecological interaction between photoautotrophic and heterotrophic species is based on cross-feeding and metabolite exchange. To date, multiprotein complexes that cross cell membrane(s) and extracellular vesicles have been evaluated in photoautotrophs for transporting materials from the interior to the exterior of the cell, which facilitate the secretion of various chemicals ranging from targeted photosynthetic intermediates, such as glycolate, osmolytes and fatty acids, and extracellular polymeric substances, to the products of cell lysis, including sugars, proteins, lipids and nucleic acids. These organic compounds could support the cell growth of the heterotrophic species [1].
The research driving this paper defines the metabolic efficiency which results when cyanobacterium Synechococcus 7942 cscB+ is co-cultured with E. coli engineered to produce 3-HP acid. The S. 7942 cscB+ strain was produced in a prior study proving that the wild type S.7942 could be engineered to export sucrose across its cellular membrane and support growth of other heterotrophic organanisms. This study confirmed the enhanced performance of both S. 7942 cscB+ and E. Coli 3-HP in a co-culture system through comparative analysis. Further, it demonstrated the consortium relieved oxidative stress and increased CO₂ availability improving the cell growth of cyanobacteria [1]. After reviewing this research we concluded it may be possible to reproduce similar results by engineering E. coli to produce chromoproteins instead of 3-HP acid.
PAPER #2:Engineering Cynaobacterium To Synthesize and Export Hydrophylic Products
Metabolic engineering of cyanobacteria has the advantage that sunlight and CO₂ are the sole source of energy and carbon for these organisms. However, as photoautotrophs, cyanobacteria generally lack transporters to move hydrophilic primary metabolites across membranes. To address whether cyanobacteria could be engineered to produce and secrete organic primary metabolites, Synechococcus elongatus PCC7942 was engineered to express genes encoding an invertase and a glucose facilitator, which mediated secretion of glucose and fructose [2].
This paper revealed some of the initial research behind the co-culture process, as well as the original patent (which has expired) confirming the results of the study. It introduced us to the S. 7942 genome, and its neutral sites 1, 2 and 3, which allow for the introduction of heterologous genes without impacting the genomic functionality of the cell [2]. The paper included the plasmid backbone, DS21, which was used to introduce the invA and glf genes from Z. mobilis as well as lacI with an IPTG regulated promoter [2]. This modification allows S. 7942 to convert CO2 into glucose and fructose from sucrose, and support their diffusion across the cell membrane. An important part of the process is an external stress which is placed on cyanobateria cells by introducing salt. This causes the cell to increase sucrose production, which helps balance osmotic pressure and maintain hydration. By engineering cyanobacteria to export glucose and fructose through a symporter protein under salt stress, the system will provide E. coli with a source of carbon for growth.
PAPER #3:Rerouting Carbon Flux To Enhance Photosynthetic Productivity
We demonstrate the efficient production of sucrose from a cyanobacterial species, Synechococcus elongatus, heterologously expressing a symporter of protons and sucrose (cscB).
In the second paper S.7942 was modified to export sucrose to E. coli using invA, which cleaves sucrose into glucose and frustose, and glf which tranports those products across the cell membrane. However, E. coli can directly uptake sucrose as a carbon source which renders invA superfluous as part of the cellular machinery. In effort to streamline and improve the efficiecy of sucrose production in S.7942, this research study introduces a sucrose permease symporter, cscB, into the S. 7942 genome through homologous recombination . The study compared the efficiency of the two variants by knocking out invA and glf, and measuring exportation of sugar molecules between them. S.7942 cscB+ exported 28% more sugar than its predecessor.
Further more, the study points out that “this productivity is the highest photosynthetic rate of microbial target product formation reported in academic literature for cyanobacteria or algae, and it exceeds (by ∼30-fold) that of a previously patented strategy for extracting sucrose from cyanobacteria” [3]. This is significant when considering how cynabacteria could revolutionize the biochemical production of chemicals for industry, especially since its carbon source is atmospheric CO₂. Given this advancement, cyanobacteria could compete with other fuel sources, and do so in an environmentally beneficial way.
SECTION 4: EXPERIMENTAL DESIGN, TECHNIQUES, TOOLS, AND TECHNOLOGY
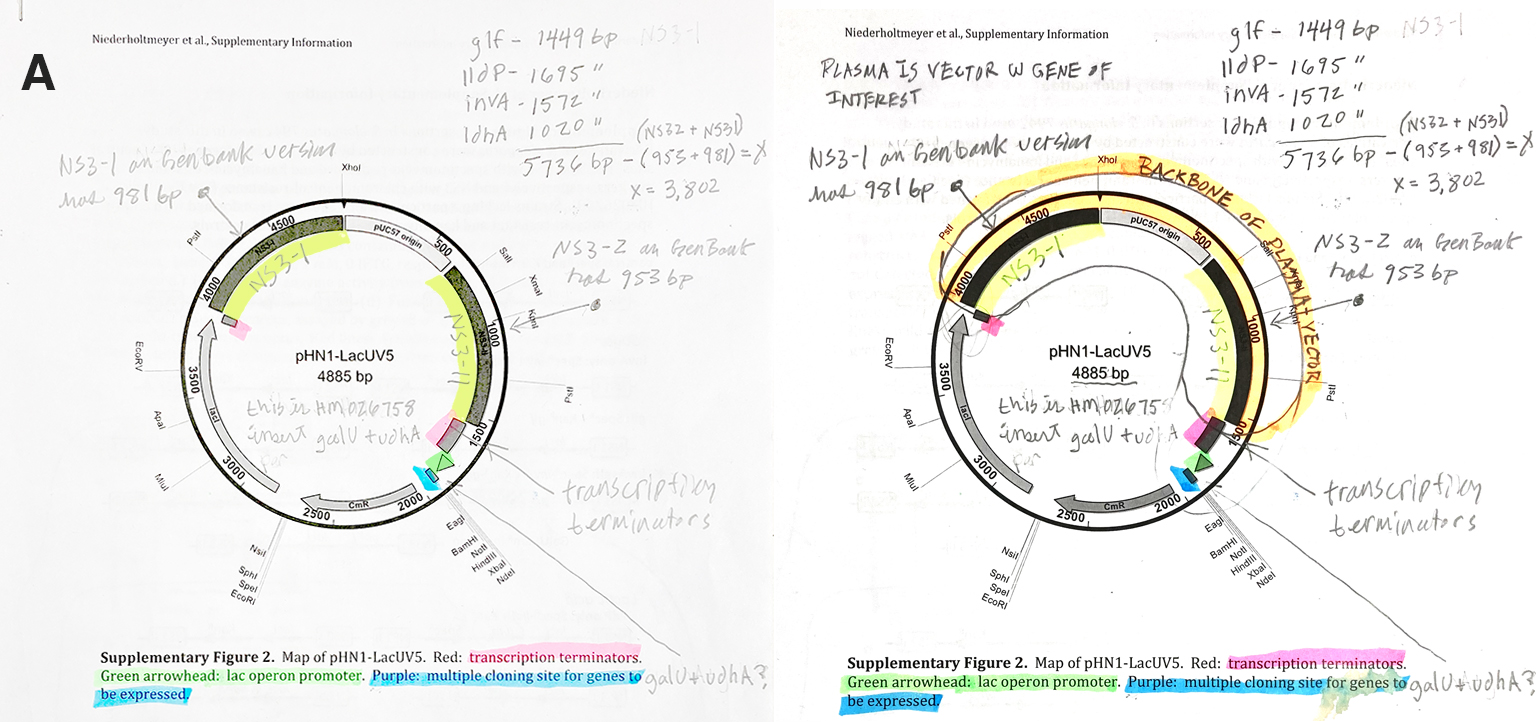
Subaim 1.1. Plasma Design The concept of plasma design is challenging, especially when considering how the collective sum of its many components determine the functionality of the plasmid relative to its host. The following image (Fig. 4) comes from my notes on the second paper, which is referenced by Cross-feeding between cyanobacterium Synechococcus and Escherichia coli. It illustrates the progression of learning plasmid theory, design, and function.
Figure 4. Plasma from article 2 (supplementary material). On the right, grasping concepts of a plasma backbone/vector.
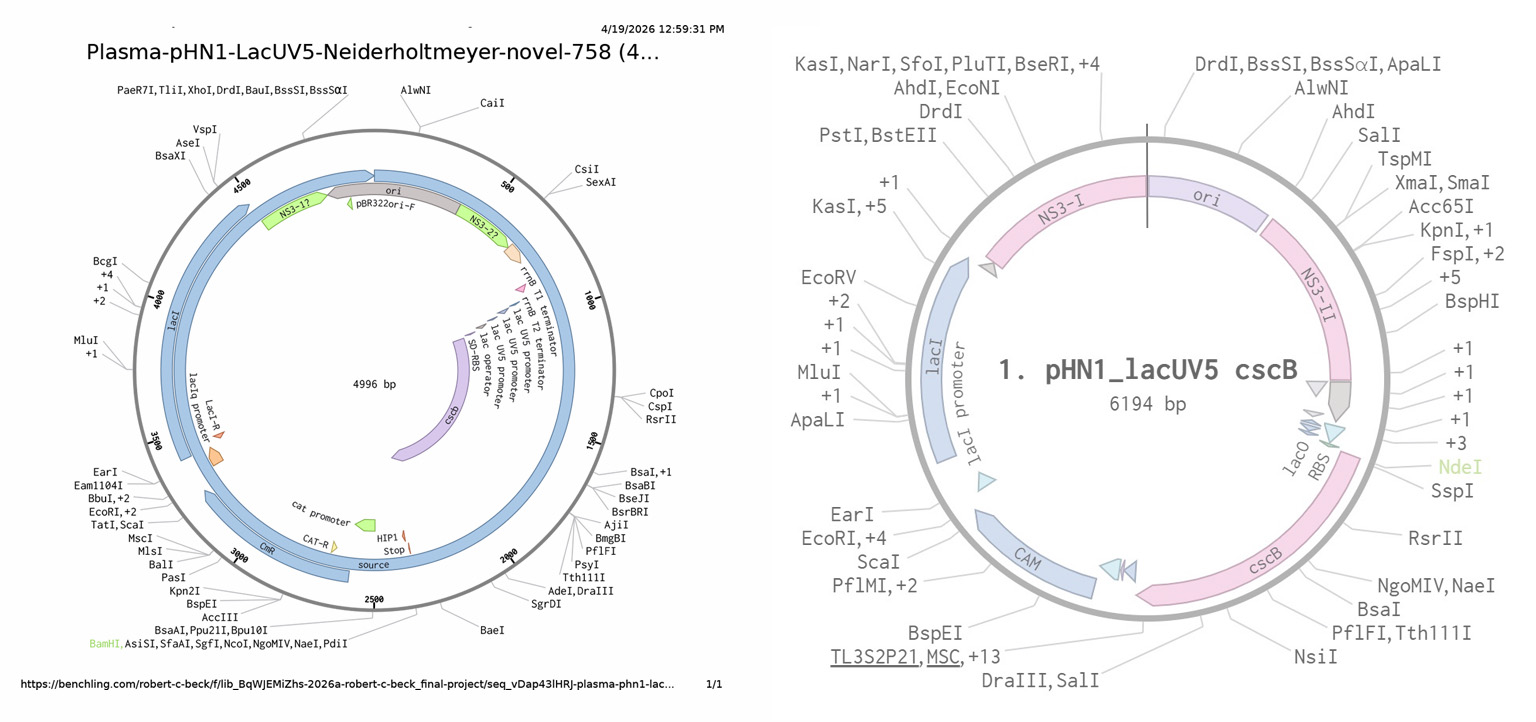
The next steps were to replicate the design in Benchling, which further challenged my understanding of synthetic biology as a practice. In the image (Fig.5) on the left hand side is my first Benchling construct for the project. Originally, when I inserted the cscB into the plasmid it pushed the number of base pairs to over 6000, which presents a problem as it relates to Twist. I thought “nuetral” sites could be substituted for functional genes since they have no effect on the target organism. My TA, Juan Diego Unda, made the functionality of the nuetral sites clear by explaining they are a neccessary component of the plasmid for it to be recognized by the target DNA and its positioning within the sequence. Figure 5. From the left, first plasma design attempt. Then with TA, Juan Diego Unda, we corrected the design (right).
Subaim 1.2. Construction of Model Organism S.7942 cscB+ (Protocol from: Specialized techniques for site-directed mutagenesis in cyanobacteria [3].)
Plasmid Construction for Mutagenesis by Homologous Recombination
Using standard molecular biology techniques (8), clone the cyanobacterial sequence
to be mutated into an E. coli cloning vector, such as pUC18 or pBR322 (we will be using pHN1-LacUV5).
Clone an antibiotic-resistance cassette within the sequence homologous to the
cyanobacterial chromosome, making sure to leave at least 300 bp of homologous
DNA flanking either side of the resistance gene for efficient recombination.
Synechococcus Transformation
Grow 100 mL of the S. elongatus strain to be mutated in liquid BG-11M to an
OD750 of 0.7.
Harvest 15 mL of cyanobacterial cells by centrifugation for 10 min at 6000g (see
Note 6).
Resuspend the cell pellet in 10 mL of 10 mM NaCl and harvest by centrifugation
for 10 min at 6000g.
Resuspend the cell pellet in 0.3 mL of BG-11M and transfer to a microcentrifuge
tube (see Note 6).
To each 0.3 mL of cells, add between 50 ng and 2 μg (typically, we use 1–2 μL
from a preparation of 100–200 ng/μL) of the recombinant plasmid that carries
the mutagenized cyanobacterial gene.
Wrap the tubes in aluminum foil to shield the cells from light and incubate them
overnight at 30°C with gentle agitation.
Plate the entire 0.3-mL cell suspension on a BG-11M plate containing the appro-
priate selective medium (see Note 2).
Incubate the plates at 30°C in constant light for approx 5 d until single colonies
appear.
Restreak isolated colonies that have the appropriate phenotypes, maintaining the
selection to favor complete segregation of mutant cyanobacterial chromosomes.
Grow mutant clones in 100 mL of BG-11M with the appropriate antibiotic to an
OD750 of 0.7. Extract the chromosomal DNA (see Subheading 3.1.3.) and verify
the presence of the mutation and its segregation on the cyanobacterial chromosome
by PCR, restriction enzyme analysis, or other technique (see Notes 4 and 7).
Extraction of Chromosomal DNA From S. elongatus
Pellet approx 10 mL of a liquid culture of cyanobacteria or scrape cells from a
plate.
Resuspend the pellet in 500 μL of 120 mM NaCl and 10 mM EDTA, pH 8.0, and
transfer the suspension to a microcentrifuge tube.
Re-pellet the cells and resuspend them in 340 μL of 25% sucrose, 50 mM Tris-
HCl, pH 8.0, 10 mM EDTA, pH 8.0. Add lysozyme to a final concentration of
2 mg/mL. Incubate the cell suspension for 45 min at 37°C.
Add 2 μL of proteinase K (from a 10 mg/mL stock solution) and 20 μL of 20%
sarkosyl, and vortex for 20 s. Incubate the mix at 55°C for 30 min.
Add 57 μL of 5 M NaCl and 45 μL of 10% cetyltrimethylammonium bromide in
0.7 M NaCl. Mix well and incubate for 10 min at 65°C.
Extract the suspension with 500 μL 24:1 chloroform:isoamyl alcohol.
Carefully transfer the upper aqueous phase to another tube and extract with 500 μL
of equilibrated phenol. Vortex for 20 s and spin for 10 min at 16,000g. The high
NaCl concentration may cause the phases to flip, placing the aqueous phase on
the bottom after the centrifugation step; the aqueous solution can be identified by
its pink hue.
Transfer the aqueous phase to another tube and extract with 500 μL of 24:1
chloroform:isoamyl alcohol mix. Vortex quickly and spin for 10 min at 16,000g.
Take the upper aqueous phase and precipitate the DNA by adding 2 v of 100%
ethanol. Mix by inverting the tube several times.
Spin down the DNA for 15 min at 16,000g in a microcentrifuge. Carefully remove
all the liquid (which contains significant salt) and wash the pellet with 1 mL of
70% ethanol. Spin again for 5 min and remove the ethanol from over the pellet.
Dry the pellet by leaving the tube open or by applying vacuum.
Resuspend the DNA in 50 μL of water and add 20 ng/μL of RNAse A (optional).
From this 50-μL final solution, 0.5 to 1 μL is typically used for a standard polymerase chain reaction (PCR).
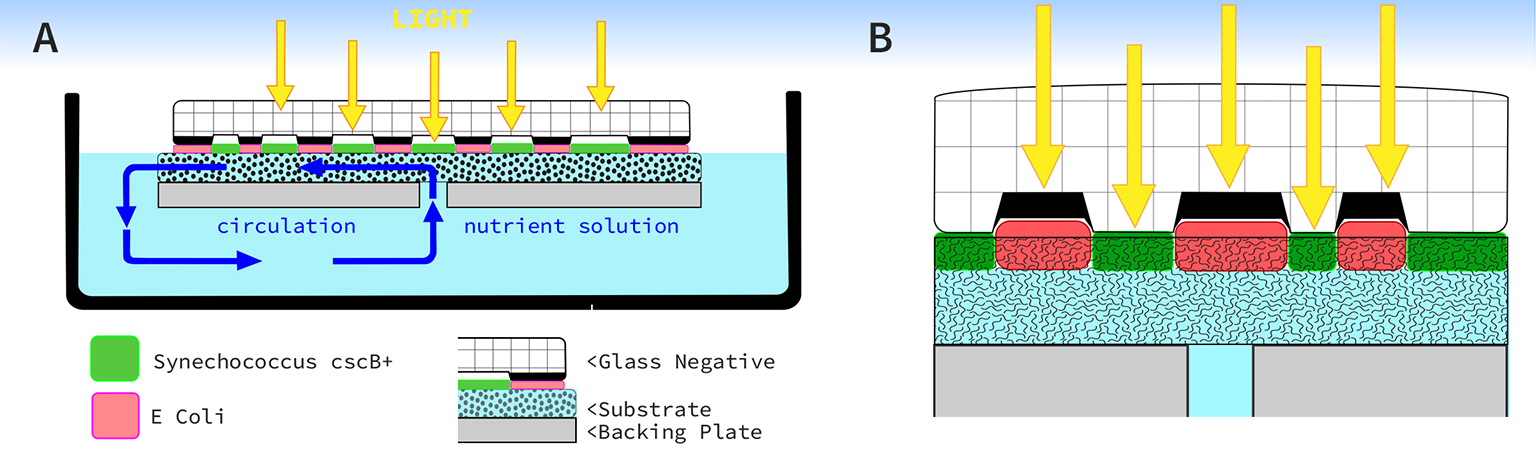
Subaim 1.3. Negative image plate design for co-culture system The original imaging system (Fig. 2) used in the 2020-2021 field experiments (Figs. 1, 3) consists of a clear glass plate with an opaque, black, raised graphic surface. The graphic is a pourous layer of low fired powdered glass which was designed to form shallow channels for cyanobacteria to grow in. After the field experiments, I noticed how the cyanobacteria grew into the substrate, and over time was over taken by another form of bacteria which grew on the surface of the substrate media (Fig 3).
My observational hypothesis is the bacteria, which took over the image, was a form of E. coli growing in the space it was afforded by the channel. If the cyanobacteria can grow into the substrate, then a new plate (Fig. 6B) could be designed to support a co-culture where engineered organisms would not compete with one another. In the illustration below (Fig. 6A), the original bio-reactor and plate design is not optimized for a co-culture system since the transparency of the channel was designed to supply light for cyanobacteria. I believe compression of the opaque areas by the raised negative graphic will inhibit E. coli formation on the substrate’s surface. The new design, reverses the spatial relationship, where opaque-negative channels provide space for chromo-protein E. coli to grow with protection from light. Adjacent to the opaque channels, clear glass will allow light to reach S. 7942 cscB+ facilitating its growth into the substrate.
Figure 6. From the left, profile of original reactor system, then redesigned image plate on right.
Aim 2. Optimization of the co-culture environment
Subaim 2.1. Test culture of S. 7942 cscB+
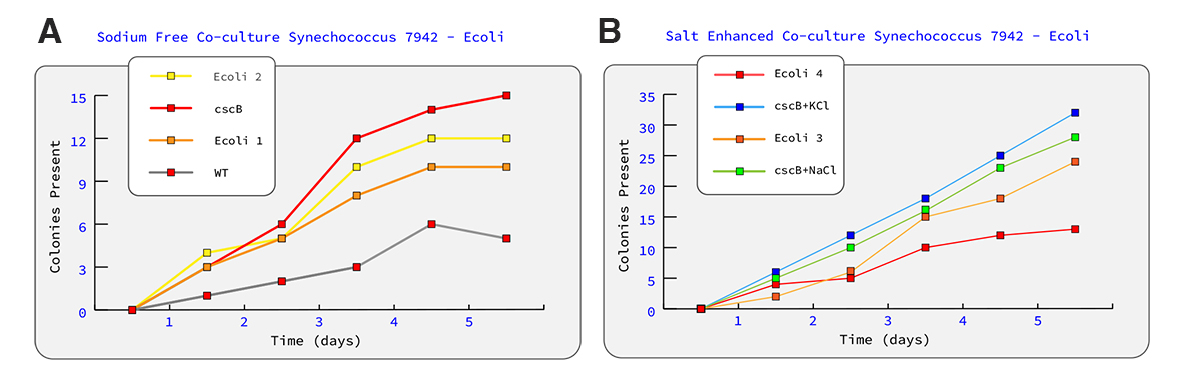
With a successful transformtion of S. 7942 into the model organism S.7942 cscB+, a co-culture medium will need to be designed along side a fluid delivery system for a recirculating bio-reactor. We will begin by optimizing the environment for culturing the model organism by following established protocols from previous research. In addition to previous research, we will test KCl as an alternate to NaCl salt (Fig. 7).
Figure 7. (Hypothetical Data). Graph A represents growth curve of sodium free cultures of PCC 7942 and E. coli expressing chromoprotein. Graph B represents two cultures which test the difference between NaCl salt induced stress in comparison with KCl as a potential alternative.
Subaim 2.3. Negative image plate fabrication Glass fabrication is expensive and energy intensive, therefore we will begin with a series of small plates designed for experiments in petri dishes (Fig. 8). If we are able to confirm the functionality of the new design, the next step (aim 3) would scale the project to accomodate larger and more complex imagery. Part of this initial research will adapt data from previous research to produce a liquid grow media formula for establishing the co-culture system in a simple petri dish bioreactor.
Figure 8. Cut glass for negative image plate tests.
Aim 3. Multicolor Systems and Mobile Technology
Subaim 3.1. Test multiple E. coli chromo-proteins within the co-culture system The possibility of generating multi color expressions with PIG is very exciting, and the ability to sustain them for extended periods of time would have a significant impact on the field of bio-art imaging. The first problem I see, past establishing a successful co-culture, is controlling the growth rate of multiple organisms within the same culture. An effective plate design may help with maintenance, but there will be other factors which will influence the system. Temperature, pH, light, CO₂, salts, and fluid dynamics all play roles in the system, and we will have to deal with them through further research and development.
Subaim 3.2. Alternative salt free systems In a seperate study exploring a different strain of Synechococcus elongatus, Utex 2973, revealed alternative engineering strategies for producing sucrose without salt stress by overexpressing sps and spp genes in addition to cscB. Without salt stress, this organism’s sucrose production was over 2-fold higher than that from the S. 7942 strain [5]. While elevated sucrose production may not be required for a chomo-protein co-culture, reducing biological stress from salt may have beneficial effects on the overall health of the system.
Subaim 3.3. Mobile lab systems for transportation of work to alternative locations While there is a considerable amount of material which needs to be researched, analysed and compiled into results, it is important to consider how the original goal of PIG, which is to produce living visual media, will function as viable form of bio-art. A portable lab for culturing imagery under a wide range of variable conditions will need to be developed. The original PIG system is in a mobile form (Fig. 9), however it will require modifications, such as the addition of a co-culture module for multi-color imagery.
Expectations Obviously, there is much research and development ahead of PIG and its future as a potential biologically controlled multi-color imagery system. Previous research with co-culturing autotrophic-heterotrophic organisms suggests the color control hypothesis is promising for bacteria based imagery. With a model organism to work with, we will be able to better define the relationships between E. coli chromo-proteins and metabolically engineered cyanobacteria.
Ethics The biotechnology driving the potential development of the project is synthetic, and must be handled in a manner applicable with the environmental laws and standards associated with handling synthetically designed organisms in any particular city, state or country. Both cyanobacteria and E. coli are water born organisms which can have significant impacts on the environment. The organsims designed for this study are lab organisms, and to our knowledge, have no modifications which could be harmful to the environment. However, as with any lab organism, they are not intended for release into the environment under any circumstances.
Further, previous research in the arena of metabolically engineered cyanobacteria reveals some very interesting characteristics these organisms can possess when they are introduced to heterologous genes. While the natural world of biology has been evolving for over four billion years, the gentic relationships we share as descendants of LUCA may hold promissing solutions to the environmental challenges many organisms face on Earth today. As humans have modified the environment, and in many cases exploited it, the potential synthetic biology holds may not only help us develop a better understanding of the environment, it could also offer many opportunities to reciprocate in a beneficial way with careful consideration.
SECTION 6: ADDITIONAL INFORMATION:
Materials and Supplies
Twist clonal gene order Q-648659
Plasmid #1: $802.88
Plasimd #2: $154.70
GeneArt® Synechococcus Engineering Kit For expression of recombinant proteins in Synechococcus elongatus
Catalog Numbers A14259
Kit A14259: 310.00bp
DH5α Competent Cells
EC0112: 128.65bp
Alternative Materials for recombinant transformation:
E. coli cloning vector, such as pUC18 or pBR322.
Modified BG-11 medium (BG-11M). Liquid medium (7): 1.5 g/L NaNO3, 0.039
g/L K2HPO4, 0.075 g/L MgSO4·7H2O, 0.02 g/L Na2CO3, 0.027 g/L CaCl2, 0.001
g/L EDTA, 0.012 g/L FeNH4 citrate, and 1 mL of the following microelement
solution: 2.86 g/L H3BO3, 1.81 g/L MnCl2·4H2O, 0.222 g/L ZnSO4·7H2O, 0.391 g/L
Na2MoO4, 0.079 g/L CuSO4·5H2O, and 0.0494 g/L Co(NO3)2·6H2O. Solid medium
Site-Directed Mutagenesis in Cyanobacteria 157
(1): equal volumes of twice-concentrated BG-11M liquid medium and Difco agar
solution (3% in sterile water) autoclaved separately and mixed together; add fil-
ter-sterilized Na2SO3 to 1 mM final concentration.
Antibiotics: kanamycin (Km), spectinomycin (Sp), cloramphenicol (Cm), strep-
tomycin (Sm), and gentamicin (Gm) (see Note 2).
10 mM NaCl (sterile).
120 mM NaCl.
10 mM EDTA, pH 8.0.
25% Sucrose (w/v in water; sterile).
50 mM Tris-HCl, pH 8.0.
10 mg/mL Lysozyme in water.
20% Sarkosyl (w/v in water).
10 mg/mL Proteinase K in water.
5 M NaCl.
10% Cetyltrimethylammonium bromide in 0.7 M NaCl.
24:1 Chloroform:isoamylic alcohol.
Equilibrated phenol (8).
100% Ethanol.
70% Ethanol.
10 mg/mL RNAse A in water.
Hit-and-run vector (see Subheading 3.3.).
E. coli strain carrying conjugal vector for conjugation (see Subheading 3.1.4.).
E. coli strain carrying helper vector for conjugation (see Subheading 3.1.4.).
Ma J, Guo T, Ren M, Chen L, Song X, Zhang W. Cross-feeding between cyanobacterium Synechococcus and Escherichia coli in an artificial autotrophic-heterotrophic coculture system revealed by integrated omics analysis. Biotechnol Biofuels Bioprod. 2022 Jun 22;15(1):69. doi: 10.1186/s13068-022-02163-5. PMID: 35733176; PMCID: PMC9219151.
Niederholtmeyer H, Wolfstädter BT, Savage DF, Silver PA, Way JC. Engineering cyanobacteria to synthesize and export hydrophilic products. Appl Environ Microbiol. 2010 Jun;76(11):3462-6. doi: 10.1128/AEM.00202-10. Epub 2010 Apr 2. Erratum in: Appl Environ Microbiol. 2010 Sep;76(17):6023. PMID: 20363793; PMCID: PMC2876443.
Ducat DC, Avelar-Rivas JA, Way JC, Silver PA. Rerouting carbon flux to enhance photosynthetic productivity. Appl Environ Microbiol. 2012 Apr;78(8):2660-8. doi: 10.1128/AEM.07901-11. Epub 2012 Feb 3. PMID: 22307292; PMCID: PMC3318813.
Clerico EM, Ditty JL, Golden SS. Specialized techniques for site-directed mutagenesis in cyanobacteria. Methods Mol Biol. 2007;362:155-71. doi: 10.1007/978-1-59745-257-1_11. PMID: 17417008.
Lin, PC., Zhang, F. & Pakrasi, H.B. Enhanced production of sucrose in the fast-growing cyanobacterium Synechococcus elongatus UTEX 2973. Sci Rep 10, 390 (2020). https://doi.org/10.1038/s41598-019-57319-5