Individual Final Project
Initially my idea was to try and connect two-ends of a chromosome (the telomeres) so that the phenomenon of telomere attrition does not happen during cell-divisions; I felt this was a good way to tackle one of the hallmarks of ageing. However, since humans have linear DNA, the problem of catenane formation is generally not encountered; due to this humans dont have the DNA topoisomerase IV (Topo IV) which helps separate the catenated chromosomes, by breaking DNA strands of one chromosome and allowing the other chromosome to pass through the break. (Thus, the replication of multiple circular DNA will have this issue of one circle passing through the other and if these circular chromosomes are being pulled to different daughter cells, then the Topo IV activity will be a must.)
The general rule [1] seems that circular chromosome organisms need dedicated decatenation machinery per chromosome: Human cells don’t have this machinery at scale, which is the fundamental problem for the circularization-idea (alongwith the additional problems of what other mutations creep in due to the Topo IV activity of breaking and making the double-stranded DNA). This idea is thus discarded!
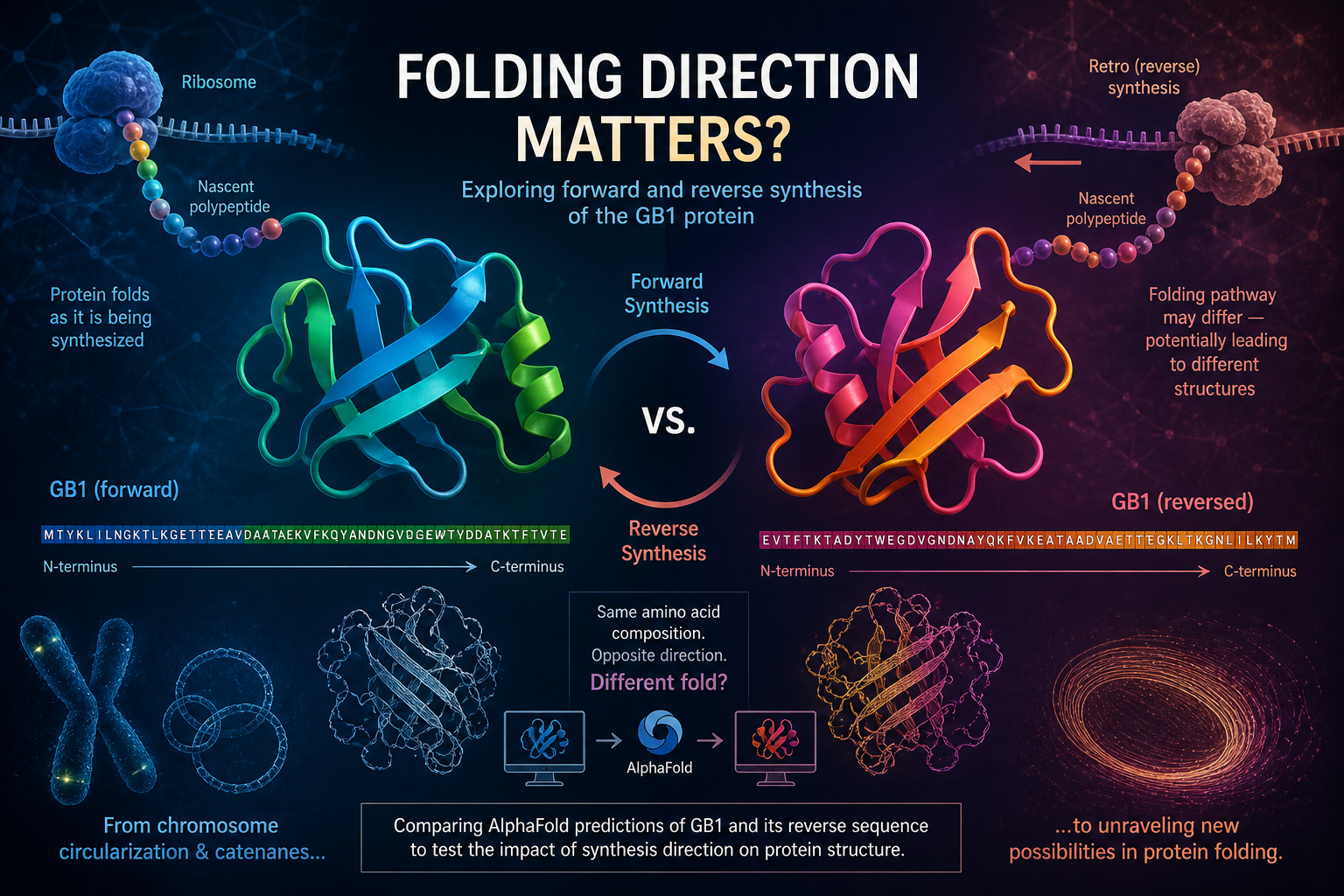
I am therefore pivoting to another idea related to protein foldings and improving AlphaFold accuracy, where I wish to understand: “How does the protein synthesis direction change the folded protein’s structure and function for the same AA-sequence?”
I have a hunch that the same AminoAcid Sequence when synthesized in the opposite direction can have drastically different folds. This is due to the fact that the rate of protein synthesis is slower than the rate of protein folding, and so the protein does not wait to be folded after it has been synthesized entirely but keeps folding on the go. Some pionees in the field suggested that these could be mirror images of the protein structure that is generated by the forward directional synthesis (Claude also thinks this is possible for some proteins, although it says that the opposite coils are not found in nature; hope I am not getting into mirror life intricacies and safety issues through this). Thus, for this experiment, I wish to find the structure of a well-known GB1 protein and its retro (reversed version), and compare these folding predictions with Alphafold’s predictions.
Final UPDATE (before website freeze): Please find my final presented slides here
Do connect if you are interested to collaborate on retro-protein foldings or global-warming induced changes in internal homeostatic/thermal landscapes leading to changes in the protein folds and potentially in the final structure.
The question to ask here is whats the maximum temperature exposure when internally there will be a lot of protein folding errors within the organism for it to sustain itself…
References
- "... Several groups of bacteria, including Brucella, Paracoccus denitrificans, and Vibrio have multiple circular chromosomes...", Conversations with Anthropic's Claude AI. https://claude.ai/share/e7198b90-5018-4c74-b60b-6b03d6c10f6a