Aim 1 Process
MycoBoard
MycoBoard engineers the fungus Neurospora crassa to grow flat hyphal mats that function as biodegradable, breadboard-like electronic substrates. Overexpression of the metallothionein gene cmt drives copper ion capture and reduction into conductive nanoparticles along hyphal walls, making the fungus an active circuit architect of the object itself. Mats grown in molds guide the hyphal geometry via its thigmotropism, and conductive tracks are applied via stamp or screen print to emulate the typical Breadboard. The result decomposes in soil within weeks or months, unlike conventional FR4 fiberglass PCBs that contribute to the ongoing e-waste crisis.
Methallothionein proteins (such as cmt) are products of mRNA translation, have a low molecular weight, and are cysteine-rich, metal-binding proteins (Cobbett & Goldsbrough, 2002). N.crassa in particular has high absorption of heavy metals because of this metallothionein protein. When the fungi is grown in heavy metal environments, or even expressing the gene in E.coli, the absorption is successful, which could demonstrate a potential for using metallothionein-based biosorption for heavy-metal bioremediation in contaminated environments (Pazirandeh et al., 1995). Additionally, “fungal mediated green chemistry approach towards the fabrication of NPs has many advantages. This includes easy and simple scale up method, economic viability, easy downstream processing and biomass handling, and recovery of large surface area with optimum growth of mycelia.” (Varshney et al., 2012)
Aim 1 - Experimental
Design and computationally validate the cmt overexpression construct for N. crassa, simulating expected copper biosorption and resistance outcomes.
To complete Aim 1 for this project, I did the following steps:
Step 1
Create a Benchling Folder: To organize all sequences and assembly data
Step 2
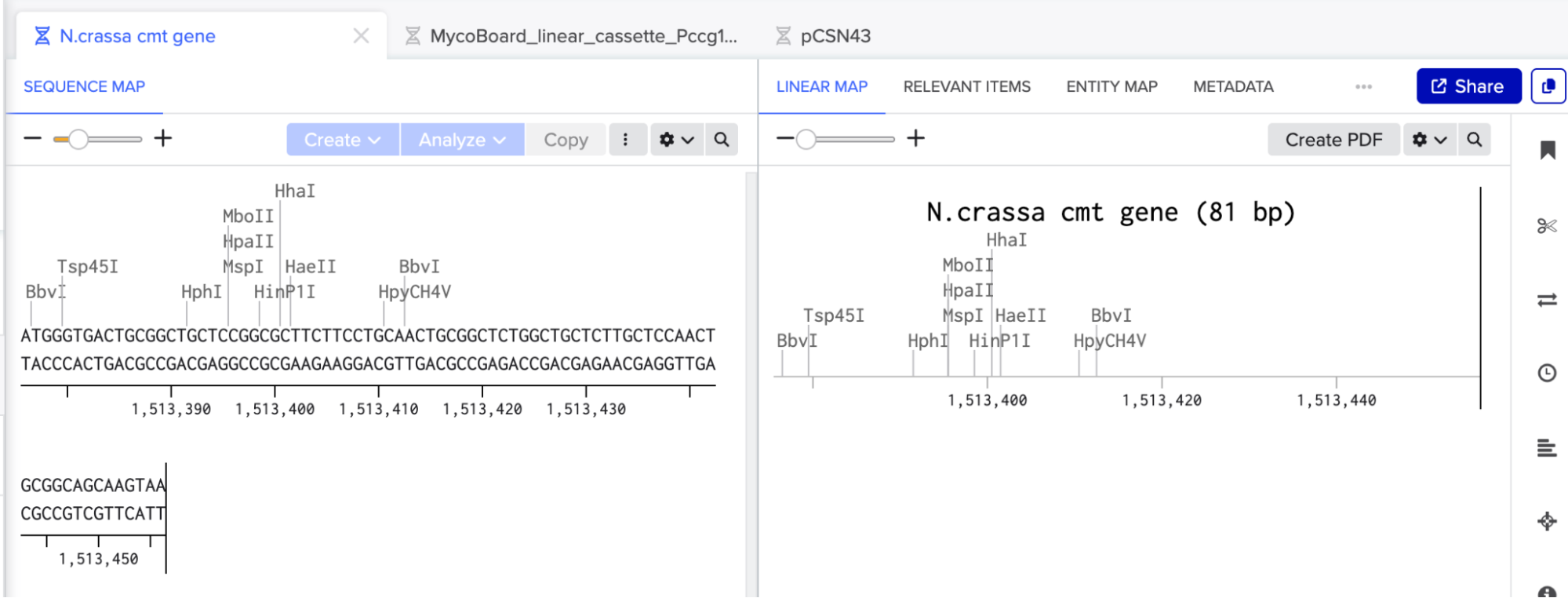
I went to FungiDB, created an account, located the Neurospora crassa cmt gene (gene ID: NCU05561), and imported its coding sequence (CDS) into Benchling.
For MycoBoard, I am using cmt to capture and reduce copper/silver ions into conductive nanoparticles along the walls of the hypha in N.crassa.
Step 3
I created a new Benchling file named MycoBoard_linear_cassette_Pccg1_cmt_HA_hygR. This file will contain the full, linear DNA construct for direct transformation into N. crassa.
N. crassa can be efficiently transformed with linear DNA via spheroplast electroporation. Fungi do not require a circular plasmid backbone for integration (like bacteria do), so the linear fragments recombine into the genome via non-homologous end joining (NHEJ) or homologous recombination. Designing the construct as a linear cassette simplifies the final transformation protocol and avoids unnecessary vector sequences, which could also make the TWIST order more expensive than it needs to be.
Step 4
I searched FungiDB for the ccg-1 gene (NCU03753). Before downloading the FASTA sequence, I adjusted the settings to obtain -1500 nucleotides upstream of the start codon (to also ensure that all regulatory elements are there, such as TATA box or transcription factor binding sites). This gave me the 1500 bp promoter region and the ccg-1 gene itself. I extracted only the promoter portion (the sequence before the ATG start codon of ccg-1) and pasted it into the linear cassette on Benchling.
Promoter sequence used (first 500 bp shown):
ACAGATGTGAAGGTATTAATTGGCAGAGAGCCGGGTCAATTCCTAGCTAGAACGGACCAG
I annotated this entire 1500 bp region as Pccg-1 promoter.
Step 5
I pasted the full N. crassa cmt CDS (84 bp) immediately after the promoter sequence. Then, I deleted the native stop codon (TAA) from the end of cmt. Immediately before where the stop codon had been, I pasted the HA tag sequence (TACCCATACGATGTTCCAGATTACGCT, 27 bp) and annotated it separately with another color. Then added a new stop codon (TGA) after the HA tag.
To create a C-terminal fusion between cmt and the HA tag, the tag must be translated as part of the same protein. Deleting the native stop codon can let the ribosome to continue translating through the HA tag before reaching the new stop codon.
The HA tag (YPYDVPDYA) allows easy detection of the cmt protein via Western blot or immunofluorescence. This is important for future wet-lab procedures to verify that the cmt protein is expressed and localized correctly in the transformed N. crassa.
I chose a C-terminal tag because the N-terminus of metallothioneins contains critical cysteine residues required for metal binding. Adding a tag to the C terminus is less likely to disrupt copper/silver ion capture.
Final cmt-HA fusion sequence (108 bp):
ATGGGTGACTGCGGCTGCTCCGGCGCTTCTTCCTGCAACTGCGGCTCTGGCTGCTCTTGCTCCAACTGCGGCAGCAAGTACCCATACGATGTTCCAGATTACGCTTGA
Translation:
M G D C G C S G A S S C N C G S G C S C S N C G S K Y P Y D V P D Y A *
Step 6
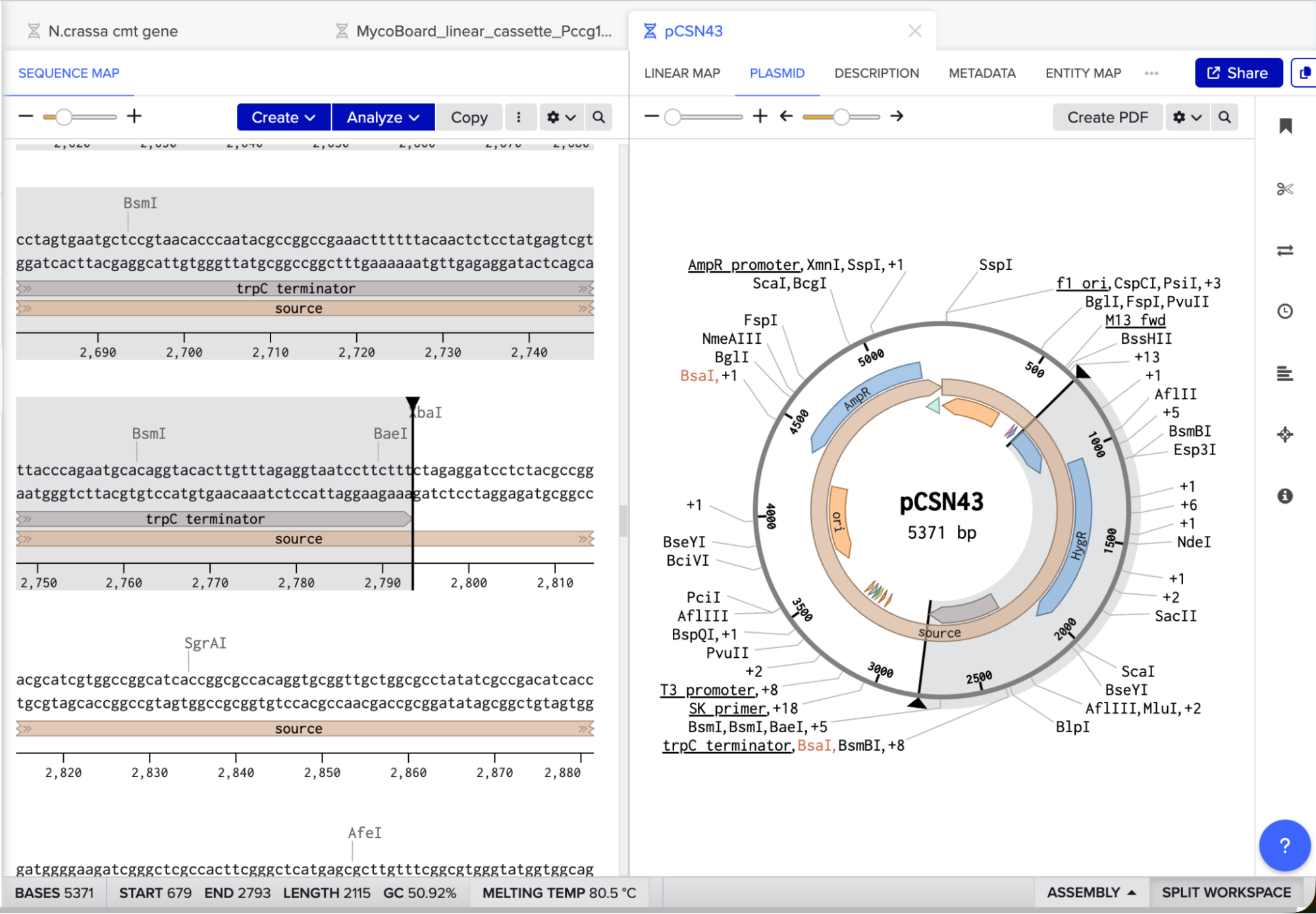
I searched for the pCSN43 vector and downloaded its GenBank file from NovoPro. I opened the file in Benchling, identified the PtrpC promoter, hygR gene, and TtrpC terminator, and copied the entire region from the start of the promoter to the end of the terminator. I pasted this cassette at the end of my linear construct (after the cmt-HA stop codon) and added a final stop codon (TAA) at the end of the hygR gene.
pCSN43 is a well-documented, widely used vector for fungal transformation. Its hygromycin B resistance cassette contains three essential parts:
- PtrpC promoter (from Aspergillus nidulans): Drives high-level expression of the resistance gene in N. crassa.
- hygR gene (hygromycin B phosphotransferase): Inactivates hygromycin B by phosphorylation, allowing transformed cells to survive on selective media.
- TtrpC terminator: Ensures proper termination of transcription and mRNA stability.
Only a small fraction of N. crassa spheroplasts will take up the engineered linear DNA. By including hygR, transformed cells can be selected on agar plates containing hygromycin B. Untransformed cells die, while successful transformed cells can grow and can be screened for cmt expression.
Also, the hygR gene requires a stop codon to terminate translation correctly. The pCSN43 cassette I downloaded did not include one, so I added TAA manually.
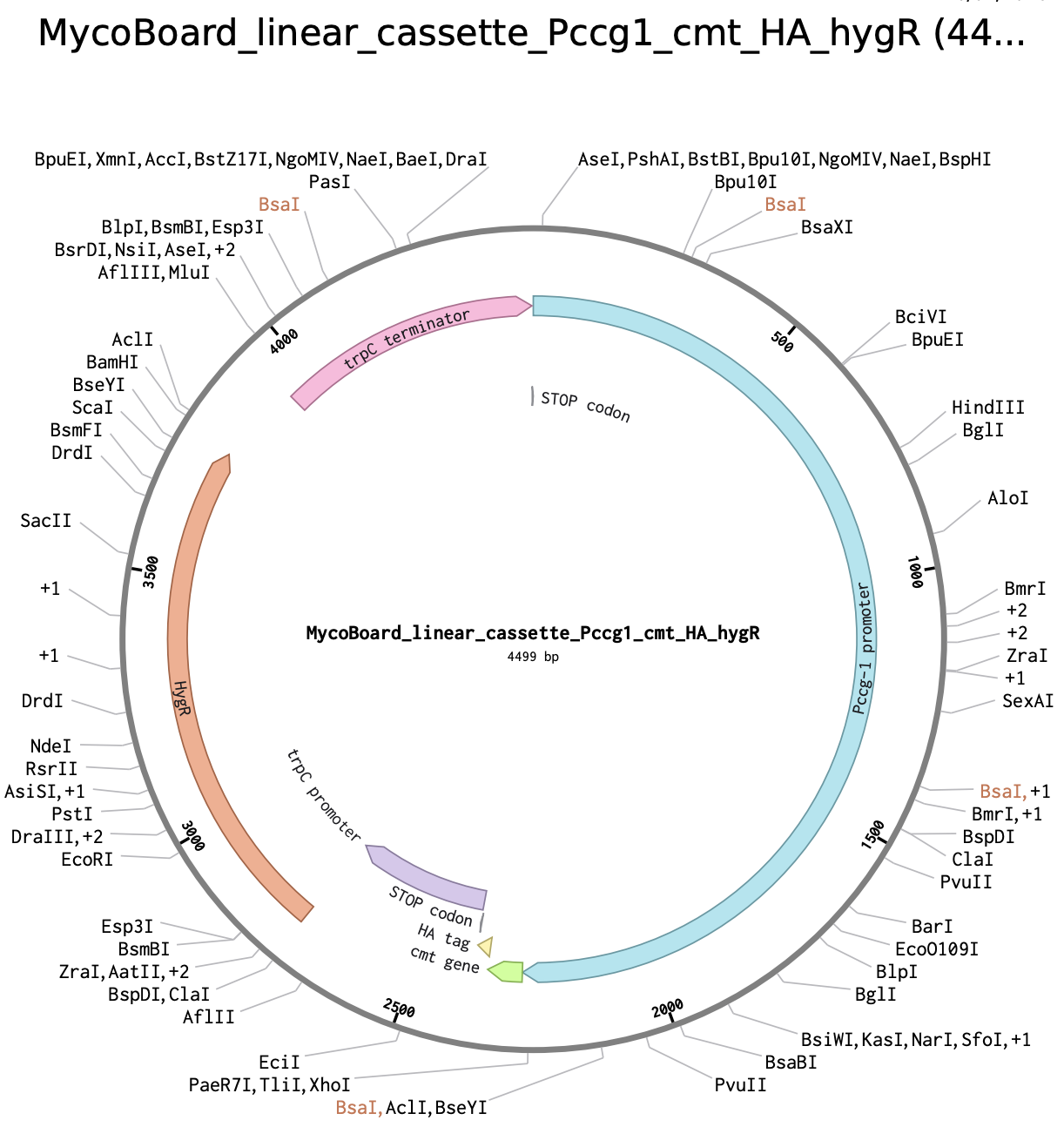
Step 7
Having the complete cassette ready in Benchling, I annotated all the elements following color-coding:
- Pccg-1 promoter (blue)
- cmt (green)
- HA tag (yellow)
- trpC promoter (purple)
- trpC terminator (pink)
- hygR (hph) (red)
- STOP codons (grey)
I then ran codon optimization on the cmt gene alone (using N. crassa as the target organism - see screenshot below for parameters) and replaced the original cmt sequence with the optimized version. Finally, I verified the reading frame by selecting the cmt and HA tag annotations together and confirming that the protein translation showed …CGSKYPYDVPDYA*.
Codon optimization is not really key for this because N. crassa is already the native host for cmt, but it could improve expression levels by matching the DNA sequence to the most abundant tRNA pools inside the organism, which is useful to express genes under strong promoters like Pccg-1
Benchling’s translation viewer also allowed to visually check that the tag is in-frame.
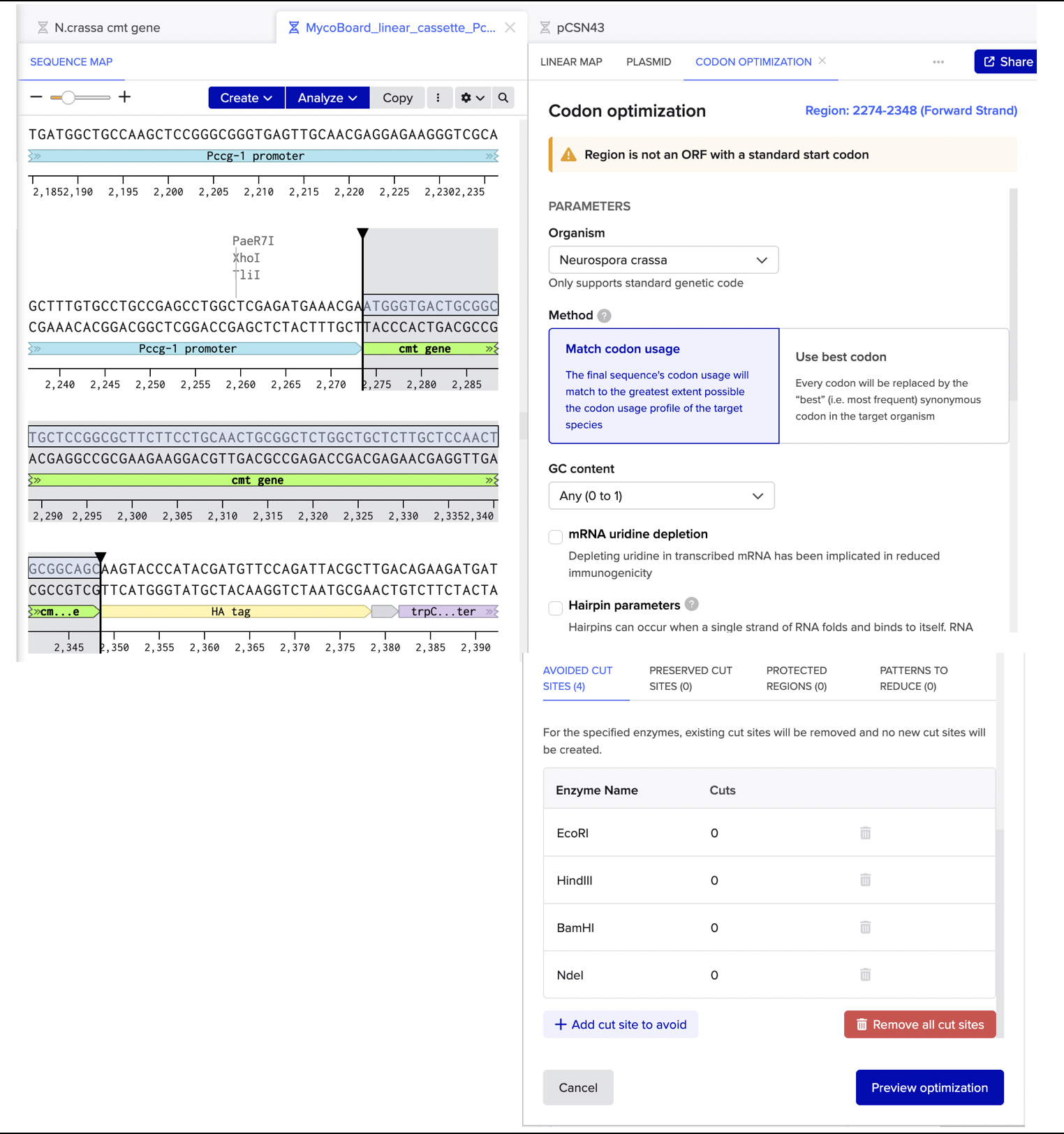
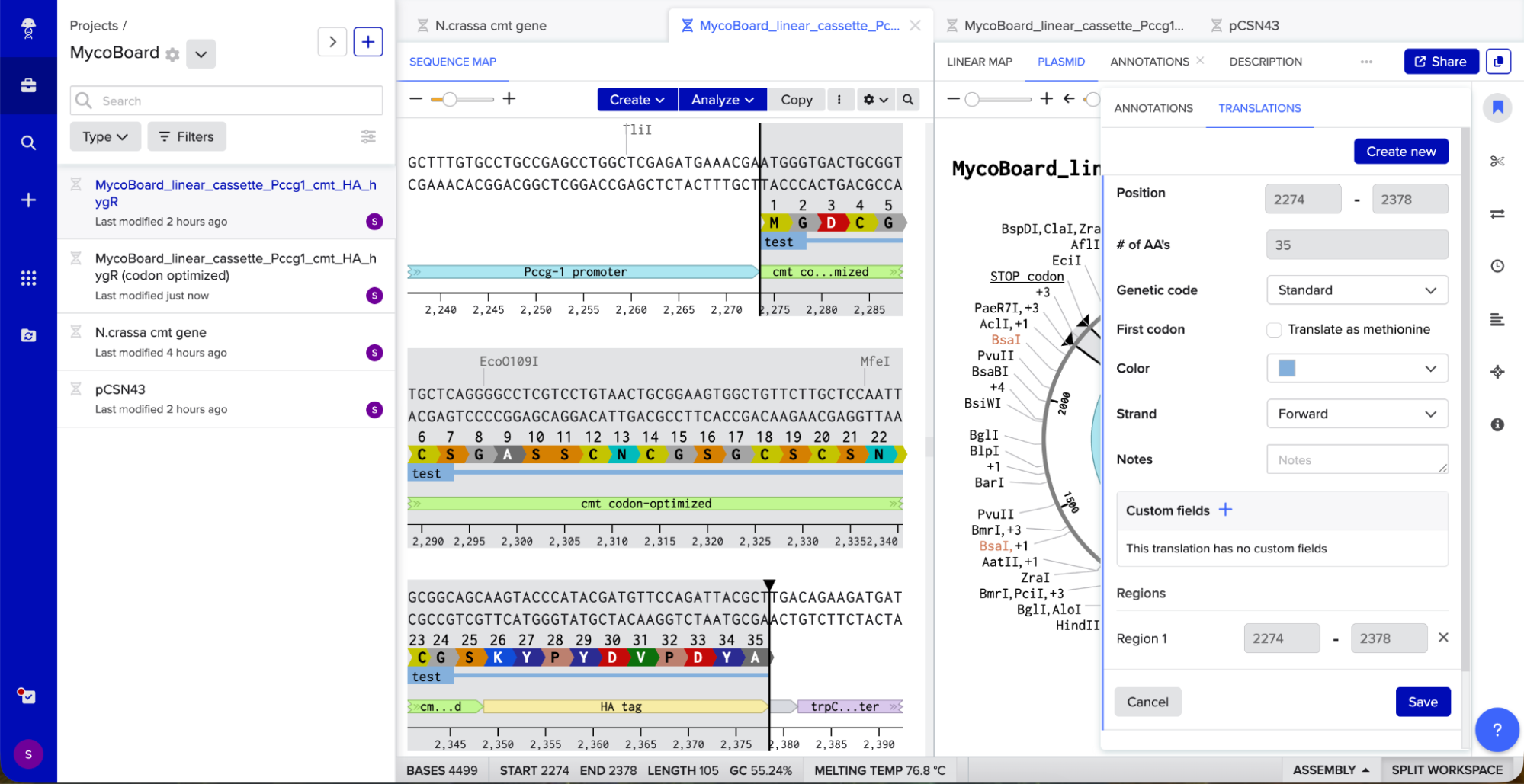
The translation looks good, as it correctly translates to the cmt protein sequence from FungiDB, the HA tag also has the correct protein translation, and there are no frame shifts. I also checked for absence of internal stop codons within the cmt coding sequence itself and there were none.
After codon optimization and reading frame verification, I prepared the Twist order. Since the sequence is longer than 1.8 kb, I decided to do two similarly sized fragments:
Fragment 1 (2,381 bp):
- Start: Beginning of Pccg-1 promoter
- End: Stop codon (TGA) after HA tag
Fragment 2 (2,118 bp):
- Start: Beginning of PtrpC promoter
- End: End of TtrpC terminator
I exported both fragments as FASTA files, went on to the Twist website and introduced the files.
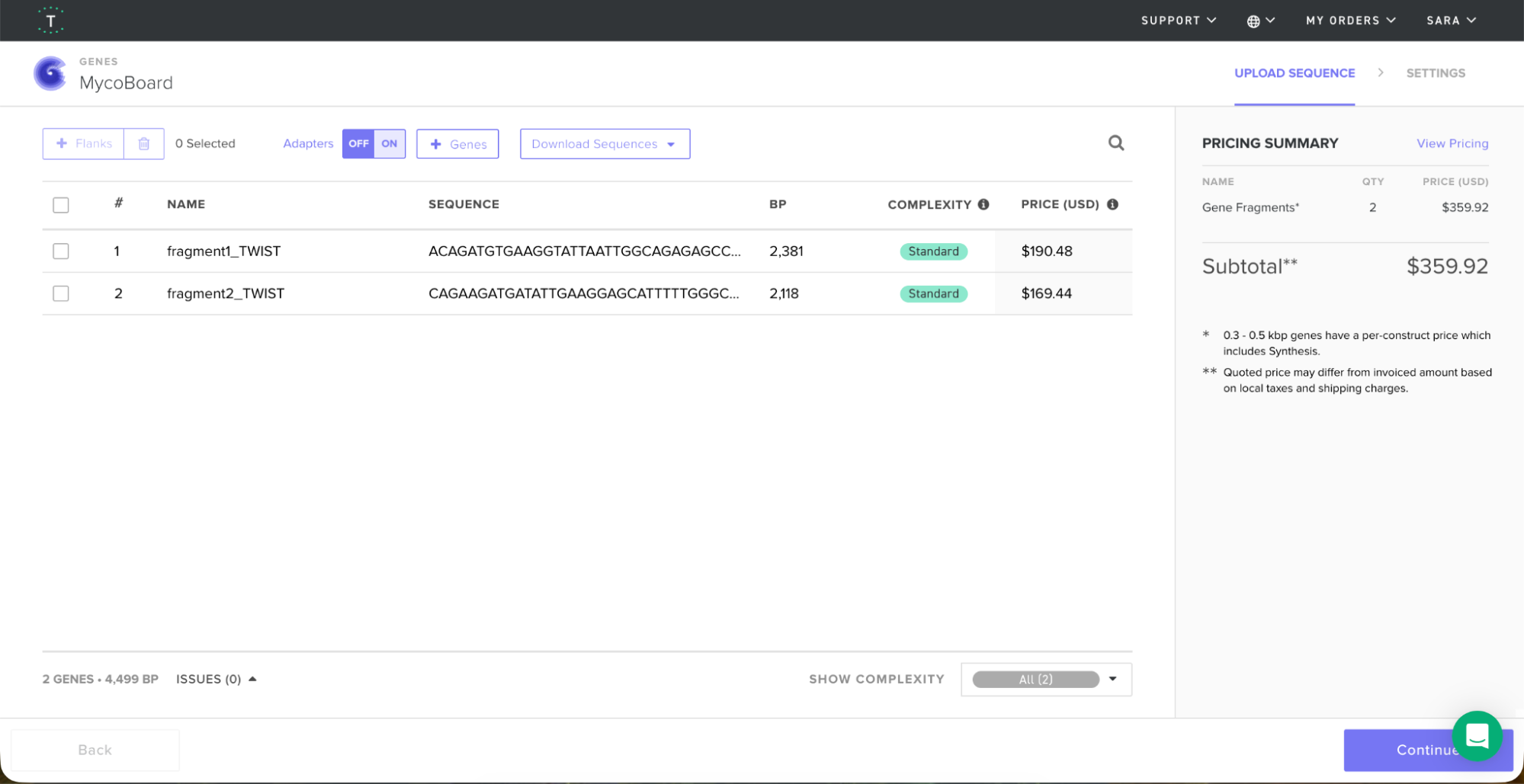
The TWIST order would come up to $359.92 for both fragments.
Validations
My original plan was to look for published data on how much silver the metallothionein protein can bind per molecule, so I could calculate the expected (or approximated) amount of silver that one square centimeter of my fungal mat could capture in experimental procedures. However, literature review revealed that while silver is highly attractive for its conductivity, N. crassa’s cmt system is much better supported as a copper-responsive metallothionein module, which makes the construct design and computational validation more feasible.
Silver still fits my broader MycoBoard vision better as a materials-oriented idea, because it is easier to imagine (and more referenced in literature) as a conductive nanoparticle track, but the fungus-specific evidence for silver capture through cmt is very limited and less straightforward.
After literature review, here is a comparison between silver and copper in relation to this project:
Silver (Ag+)
- Can act as an antifungal and antimicrobial agent (Jabran et al., 2025; Mohan et al., 2024).
- Has a higher electrical conductivity, but is more expensive (Robinson et al., 2022).
- Most fungal genera are coupled with the synthesis of silver nanoparticles intracellularly or extracellularly, and this has been widely researched and characterized (Varshney et al., 2012).
- Less research on silver capture by N. crassa
Copper
- Can act as an antifungal and antimicrobial agent (Jabran et al., 2025; Mohan et al., 2024). In one of the papers, Yoon et al. (2005) observed that copper had a superior antibacterial activity compared to the silver nanoparticles.
- Has a lower electrical conductivity, but is cheaper (Robinson et al., 2022).
- Less research available on the biological synthesis of copper nanomaterials with size and shape control (Varshney et al., 2012).
- Well described cloning of the cmt promoter from N. crassa (Ohrnberger & Akins, 1995) and understanding of the process of how N. crassa accumulates copper with copper-binding proteins such as metallothionein (Lerch, 1980), and how the presence of copper alone (without the need of oxidative stress) induce the cmt gene expression (Kumar et al., 2005).
This decision does not require changing the construct design, the Benchling cassette uses the well-documented ccg-1 constitutive promoter and the native cmt coding sequence, both of which are fully compatible with a copper-responsive validation. The validation now focuses on the theoretical copper capture capacity of the mat:
Copper Biosorption and Stoichiometry
Literature confirms that N. crassa copper accumulation is well-documented: “when N. crassa is grown in the presence of Cu(II) ions, it accumulates the metal with the concomitant synthesis of a low molecular weight copper-binding protein. The molecule binds 6 g-atom of copper per mole protein (Mr = 2200) and shows a striking sequence homology to the zinc- and cadmium-binding vertebrate metallothioneins.” (Beltramini & Lerch, 1986).
Since I couldn’t find published values for cmt abundance in a determined area of N. crassa fungal mats, such as mass of total cmt protein per cm² of fungal mat (or similar area measurement), mass of cmt per mass of total protein, or cmt expression levels from a quantitative assay like Western blot densitometry, I searched for published biosorption values for N. crassa.
Research by Suresh and Subramanyam (1998), Beltramini and Lerch (1986), Subramanyam et al. (1983), Germann and Lerch (1987), and the copper-metallothionein characterization work on N. crassa indicate that copper stress can produce blue mycelium and substantial copper accumulation in N. crassa. Key results include:
- At 0.63 mM copper, N. crassa showed about 80% growth inhibition and blue mycelium/cell-wall coloration (Suresh & Subramanyam, 1998).
- At 0.5 mg Cu per 10 mL medium, growth inhibition was about 50%, and at stronger copper stress the mycelial copper content reached about 8 mg per 100 mg dry weight (Subramanyam et al., 1983).
- The blue mycelium cell wall contained about 12% copper and had a Cu/phosphate molar ratio of 7.18 under the cited condition (Subramanyam et al., 1983).
- Copper uptake in N. crassa occurred during exponential growth, starting around 30 h after inoculation and reaching saturation by 70 h, and the wild type accumulated more copper than the slime mutant under the same CuSO4 exposure (Germann & Lerch, 1987).
- At 0.5 mM CuSO4, the slime mutant retained about 23% of the initial copper in the medium, compared with 51% for the wild type (Germann & Lerch, 1987).
- The purified N. crassa copper metallothionein binds 6 Cu per molecule and forms a Cu(I)-thiolate cluster (Beltramini & Lerch, 1986).
Theoretical Copper Loading Calculation
Using the mentioned stoichiometry, if one mole of cmt binds 6 Cu ions, then:
6 × 63.546 = 381.276 g Cu per mol cmt
The molecular mass of cmt is approximately 2,200 g/mol (Lerch, 1980), so the mass of a fully copper-loaded cmt molecule is the sum of the protein mass and the bound copper mass.
Including the approximate protein mass of cmt of 2200 g/mol gives:
2200 + 381.276 = 2581.276 g/mol
So the copper mass fraction in fully loaded Cu6-cmt is:
381.276 / 2581.276 = 0.147708
Meaning fully Cu loaded cmt is about 14.77% copper by mass.
If there are 8 mg of Cu per 100 mg dry mycelium, that is 8% copper by mass. Dividing the observed copper fraction by the copper fraction of fully loaded cmt gives:
0.08 / 0.147708 = 0.541608
So about 54.2% of the dry biomass would need to behave like fully loaded Cu6-cmt to explain the full copper content.
If the fungal mat for MycoBoard has 10 mg dry mass per cm², then using the paper’s copper level:
10 × 0.08 = 0.8 mg Cu/cm²
That is 0.8 mg Cu/cm², or about 12.59 µmol Cu/cm².
This is a validated upper-bound benchmark for the copper-loading capacity of the engineered fungal mat for MycoBoard. Even though approximately 54% of the biomass would need to be fully saturated cmt to account for this calculated copper, the actual copper pool is likely distributed between cmt, cell wall polyphenol binding (Suresh & Subramanyam, 1998), and other intracellular components.
Restriction enzyme analysis
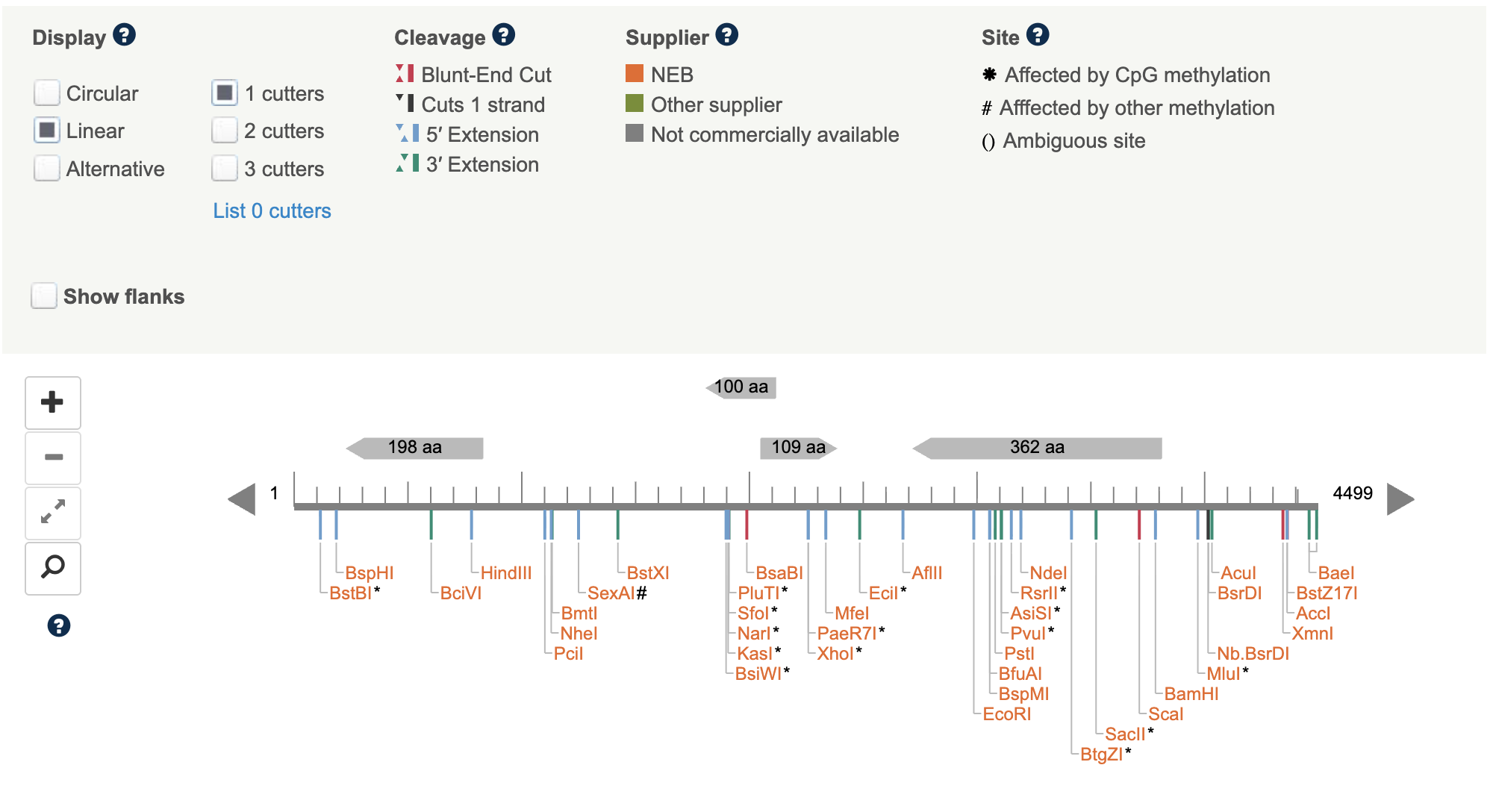
For future applications of the cassette, I decided to use NEB Cutter to identify cut sites within the linear cassette.
- No enzyme cut sites were found within the HA tag sequence.
- Two enzymes (Eco0109I and MfeI) cut within the codon-optimised cmt coding sequence, as the cassette is designed for direct linear transformation instead of restriction enzyme cloning, these sites do not affect construct function but hinder the use of these enzymes for future digestion of the cmt region.
- Multiple restriction sites were identified within the hygR resistance cassette, including some commonly used enzymes EcoRI, PstI, and NdeI. These are also consistent with the published pCSN43-derived hph sequence and expected for a ~1,000 bp coding region. These sites do not affect hygromycin resistance function but limit options for future restriction-based cloning of the cassette. Additionally from these common enzymes, four Type IIS enzyme sites (Esp3I, BsmBI, BfuAI, BspMI) were also identified within hygR, indicating that Golden Gate Assembly-based approaches would require removal of these sites before use in cloning methods.
Aim 2 - Development
Translate the validated computational design into a physical prototype: grow engineered N. crassa mats inside molds and validate by working LED circuit powered through a biologically grown conductive track.
- Transform verified construct into N. crassa via spheroplast electroporation
- Grow flat mats inside laser-cut or 3D-printed breadboard-like shaped molds
- N.crassa hyphae follow channel walls via thigmotropism, forming predefined track layout
- Soak mat in AgNO₃ medium (Copper medium if silver is not accessible), heat-fix and dry
- Map resistance across mat with multimeter to confirm conductivity along tracks
- Mount a simple LED + resistor circuit with conductive adhesive and validate conductivity
Aim 3 - Visionary
Replace physical molds with optogenetic control of cmt expression, using an interface to draw circuit layouts that the fungus then follows while growing.
- Engineer light-inducible promoter controlling cmt
- Design circuit layout and project onto growing mat. Illuminated zones activate cmt, copper deposits follow the projected pattern precisely
- Characterise mat decomposition in soil, revise chelation wash protocol if helpful
- Generalise platform to other metals and fungal chassis (perhaps other species like Fusarium or Ganoderma)
- Expected result: custom biodegradable boards grown from spores, compostable in months
REFERENCES
Nargang, C. E., & Nargang, F. E. (1996). Procedure for preparing and transforming spheroplasts of Neurospora crassa.
Singh, K., Sharma, S., Kalia, A., & Manchanda, P. (2026). Advancement in Mushroom Transformation: From Conventional Techniques to Modern Genetic Engineering. Journal of Basic Microbiology, 66(1), e70132.
Ploessl, D. (2022). Developing nuclear and mitochondrial DNA editing techniques for engineering yeasts as novel microbial factories and disease models (Doctoral dissertation, Iowa State University).
Wang, Z., Bartholomai, B. M., Loros, J. J., & Dunlap, J. C. (2023). Optimized fluorescent proteins for 4-color and photoconvertible live-cell imaging in Neurospora crassa. Fungal Genetics and Biology, 164, 103763.
Staben, C., Jensen, B., Singer, M., Pollock, J., Schechtman, M., Kinsey, J., & Selker, E. (1989). Use of a bacterial hygromycin B resistance gene as a dominant selectable marker in Neurospora crassa transformation. Fungal Genetics Reports, 36(1), 79.
Danninger, D., Pruckner, R., Holzinger, L., Koeppe, R., & Kaltenbrunner, M. (2022). MycelioTronics: Fungal mycelium skin for sustainable electronics. Science Advances, 8(45), eadd7118.
Riquelme, M., Aguirre, J., Bartnicki-García, S., Braus, G. H., Feldbrügge, M., Fleig, U., … & Fischer, R. (2018). Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiology and Molecular Biology Reviews, 82(2), 10-1128.
Rai, M., Bonde, S., Golinska, P., Trzcińska-Wencel, J., Gade, A., Abd-Elsalam, K. A., … & Ingle, A. P. (2021). Fusarium as a novel fungus for the synthesis of nanoparticles: mechanism and applications. Journal of Fungi, 7(2), 139.
Ohrnberger, J., & Akins, R. A. (1995). Cloning of the copper-inducible metallothionein (cmt) promoter from Neurospora crassa. Fungal Genetics Reports, 42(1), 57-58.
Baldé, C. P., Kuehr, R., Yamamoto, T., McDonald, R., D’Angelo, E., Althaf, S., … & Wagner, M. (2024). Global e-waste monitor 2024.
Carroll, A. M., Sweigard, J. A., & Valent, B. (1994). Improved vectors for selecting resistance to hygromycin. Fungal Genet. Newsl, 41(22), 135-143.
Romeyer, F. M., Jacobs, F. A., & Brousseau, R. (1990). Expression of a Neurospora crassa metallothionein and its variants in Escherichia coli. Applied and environmental microbiology, 56(9), 2748-2754.
Cobbett, C., & Goldsbrough, P. (2002). Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Annual review of plant biology, 53(1), 159-182.
Jabran M, Shafique MS, Abbas A, Han W, Ali MA, Gao L. Antifungal potential of silver and copper nanoparticles in suppressing Tilletia indica for Karnal bunt resistance in wheat. BMC Plant Biol. 2025 Nov 6;25(1):1517. doi: 10.1186/s12870-025-07547-x. PMID: 41199180; PMCID: PMC12590600.
Mohan, C. R., Kandasamy, R., & Kabiriyel, J. (2024). Correlation between electrical conductivity and antibacterial activity of chitosan-stabilized copper and silver nanoparticles. Carbohydrate Polymer Technologies and Applications, 7, 100503.
Robinson, J., Munagala, S. P., Arjunan, A., Simpson, N., Jones, R., Baroutaji, A., … & Lyall, I. (2022). Electrical conductivity of additively manufactured copper and silver for electrical winding applications. Materials, 15(21), 7563.
Yoon, K. Y., Byeon, J. H., Park, J. H., & Hwang, J. (2007). Susceptibility constants of Escherichia coli and Bacillus subtilis to silver and copper nanoparticles. Science of the Total Environment, 373(1), 572–575.
Varshney, R., Bhadauria, S., & Gaur, M. S. (2012). A review: biological synthesis of silver and copper nanoparticles. Nano Biomedicine & Engineering, 4(2).
Lerch, K. (1980). Copper metallothionein, a copper-binding protein from Neurospora crassa. Nature, 284(5754), 368-370.
Kumar, K. S., Dayananda, S., & Subramanyam, C. (2005). Copper alone, but not oxidative stress, induces copper–metallothionein gene in Neurospora crassa. FEMS microbiology letters, 242(1), 45-50.
Beltramini, M., & Lerch, K. (1986). Primary structure and spectroscopic studies of Neurospora copper metallothionein. Environmental health perspectives, 65, 21.
Pazirandeh, M., Chrisey, L.A., Mauro, J.M. et al. Expression of the Neurospora crassa metallothionein gene in Escherichia coli and its effect on heavy-metal uptake. Appl Microbiol Biotechnol 43, 1112–1117 (1995). https://doi.org/10.1007/BF00166934
Nielson KB, Atkin CL, Winge DR. Distinct metal-binding configurations in metallothionein. J Biol Chem. 1985 May 10;260(9):5342-50. PMID: 3988757.
Suresh, K., & Subramanyam, C. (1998). Polyphenols are involved in copper binding to cell walls of Neurospora crassa. Journal of inorganic biochemistry, 69(4), 209-215.
Subramanyam, C., Venkateswerlu, G., & Rao, S. L. N. (1983). Cell wall composition of Neurospora crassa under conditions of copper toxicity. Applied and environmental microbiology, 46(3), 585-590.
Germann, U. A., & Lerch, K. (1987). Copper accumulation in the cell-wall-deficient slime variant of Neurospora crassa. Comparison with a wild-type strain. Biochemical journal, 245(2), 479-484.
Beltramini M, Lerch K. Primary structure and spectroscopic studies of Neurospora copper metallothionein. Environ Health Perspect. 1986 Mar;65:21-7. doi: 10.1289/ehp.866521. PMID: 3011391; PMCID: PMC1474700.