HTGAA 2026: Individual Final Project Documentation
FINAL PRESENTATION SLIDES
SECTION 1: ABSTRACT
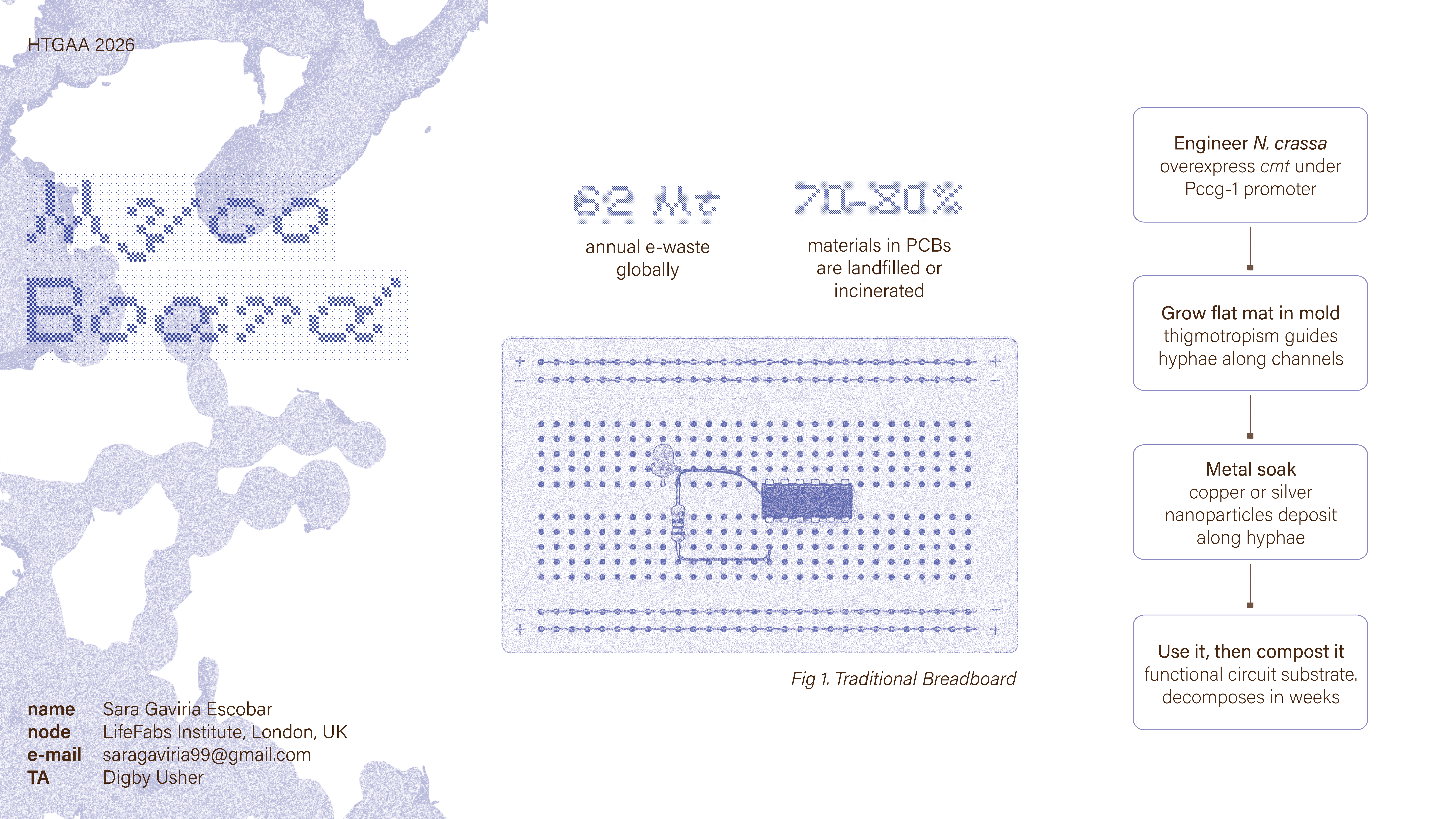
MycoBoard addresses the global e-waste crisis by engineering the fungi Neurospora crassa to grow biodegradable hyphal mats that function as breadboard-like electronic substrates. Conventional FR4 fiberglass PCBs contribute to the 62 million metric tons of annual e-waste, most of which is non-recyclable. MycoBoard leverages fungal thigmotropism to form conductive tracks within molded mats, with cmt metallothionein overexpression driving copper ion capture along the hyphae walls of N.crassa. The purpose is to create compostable electronic substrates that can break down in soil within weeks. The principle, validated by a Benchling-designed linear cassette and literature-based copper-loading estimates, states that engineered N. crassa could both biosorb copper and form conductive pathways via filamentous formation of hyphae. Aim 1 designs and validates a linear cassette construct for copper-responsive expression of cmt. Aim 2 transforms N. crassa experimentally, grows mats in breadboard molds, and tests LED circuit conductivity. Aim 3 replaces molds with optogenetic cmt control for pattern-directed copper deposition. Methods include Benchling DNA design, Twist synthesis, spheroplast electroporation, fungal mat cultivation, and multimeter/resistance validation.
SECTION 2: PROJECT AIMS
Aim 1 - Experimental Aim (executed):
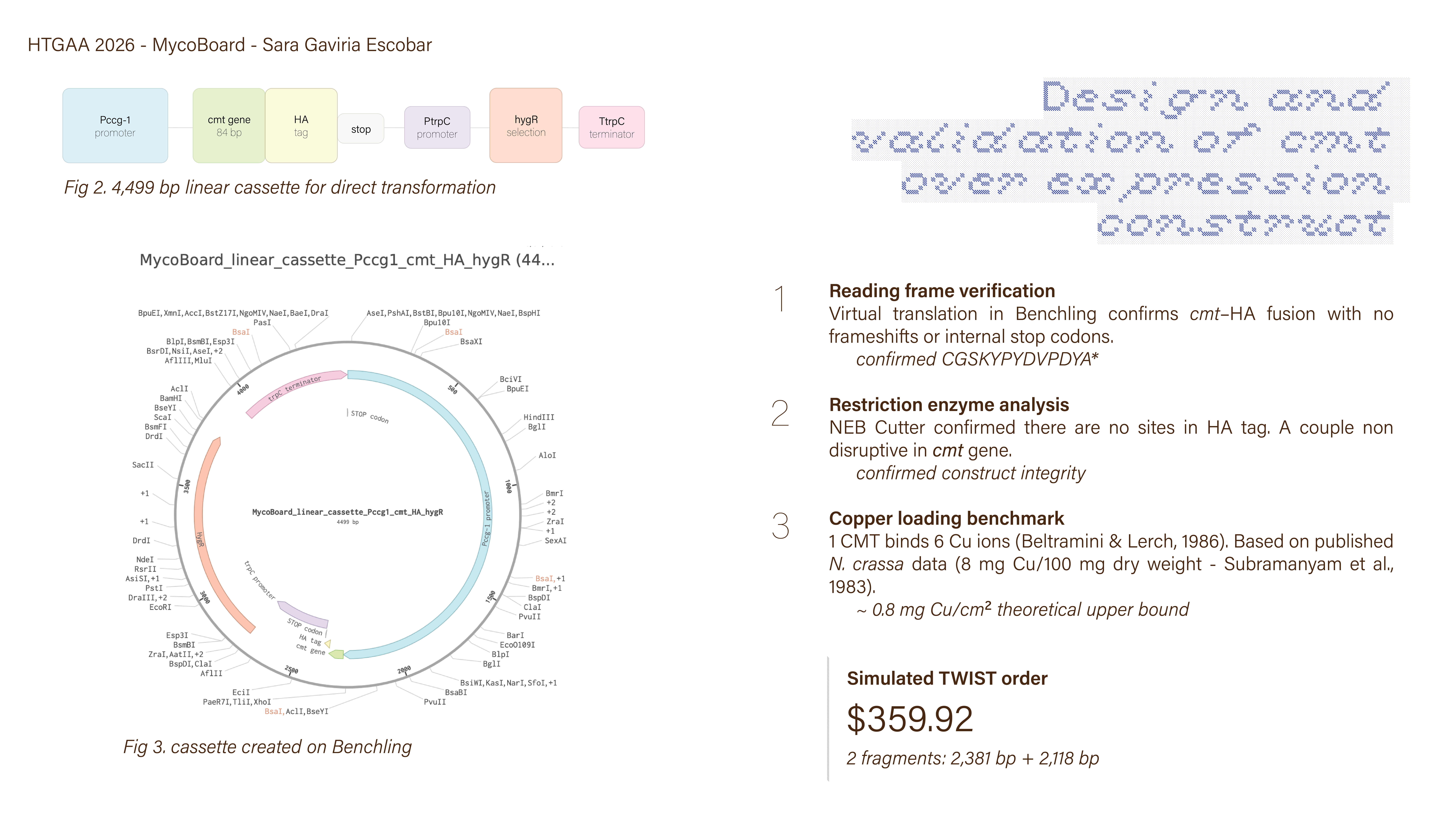
The first aim of my final project is to design and computationally validate a Pccg-1=cmt-HA=hygR linear cassette for Neurospora crassa overexpression of cmt protein, using Benchling for sequence assembly, codon optimization, HA-tag fusion verification, and Twist order preparation. Methods include FungiDB for gene and promoter search, NEB Cutter for restriction site analysis, and literature-based copper biosorption estimations and predictions by N. crassa. The outcome is a complete Benchling file with verified reading frame and theoretical copper capture benchmark data of concentration of copper in a specific area of the fungal mat.
Aim 2 - Development Aim:
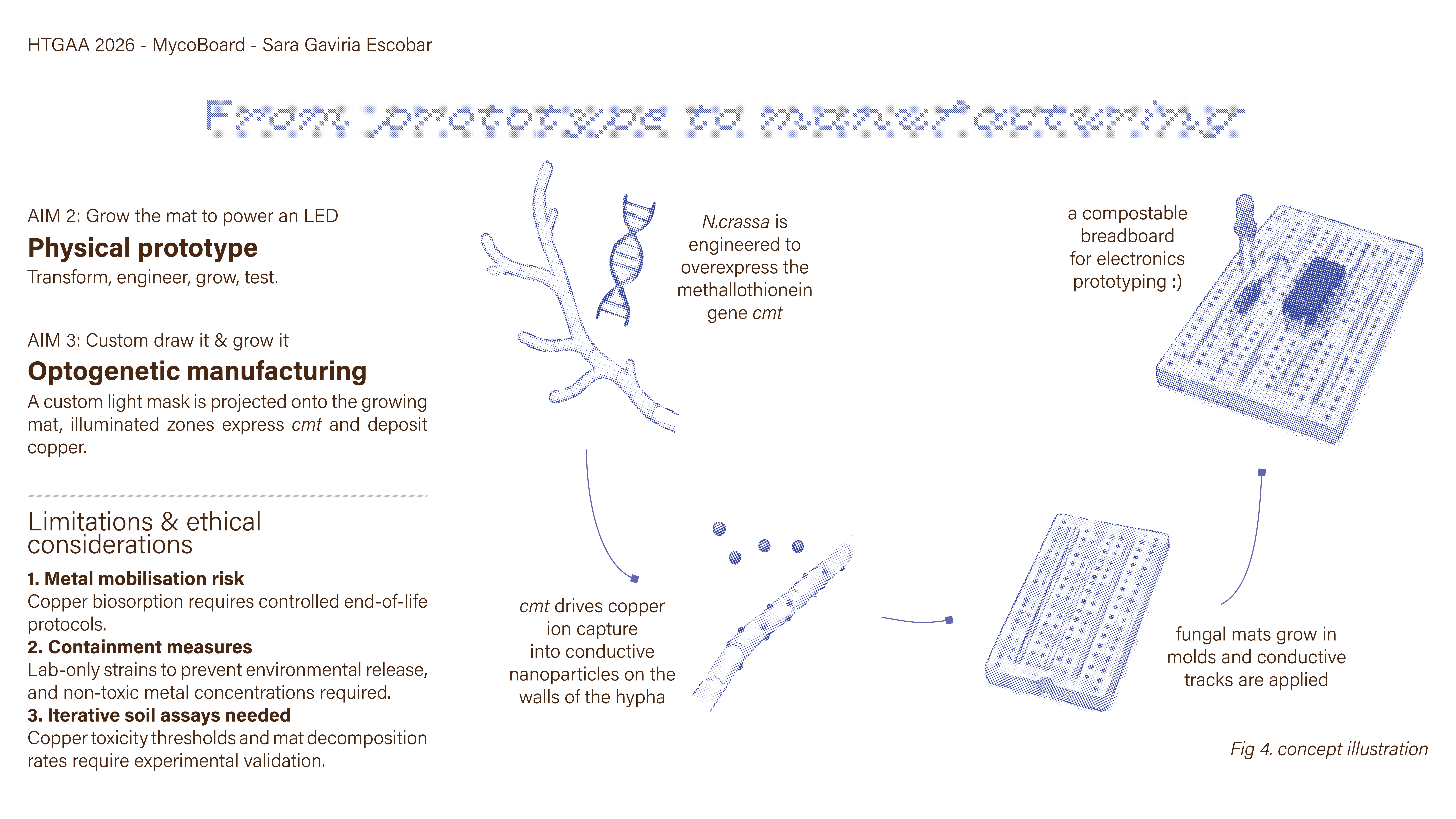
Translate the validated construct into a physical prototype by experimentally transforming N. crassa through spheroplast electroporation, then growing engineered mats in laser-cut/3D-printed breadboard-like molds, soaking in copper medium, and validating conductivity with a multimeter-powered LED circuit. This extends Aim 1 by testing hyphal thigmotropism first hand to track formation and copper deposition on the fungal mat for electrical conductivity of the material.
Aim 3 - Visionary Aim:
Replace physical molds with optogenetic control of cmt expression through a light-inducible promoter, allowing users to define custom circuit layouts that direct copper deposition during fungal growth. This realizes unique and on-demand compostable PCBs grown from spores, avoiding FR4-related pollution and enabling sustainable electronics production at scale.
SECTION 3: BACKGROUND
Beltramini & Lerch (1986) characterized Neurospora crassa copper metallothionein (cmt) as a low-molecular-weight protein binding 6 Cu(I) ions per molecule (Mr = 2200), induced by Cu(II) stress without any necessary oxidative action required. The cmt protein forms a Cu(I)-thiolate cluster homologous to vertebrate MTs, supporting metal homeostasis. Subramanyam et al. (1983) and further researchers showed copper toxicity produces blue mycelium with 8 mg Cu/100 mg dry weight and cell walls containing 12% copper, indicating successful and important copper biosorption capacity.
MycoBoard’s novelty is repurposing N. crassa’s natural copper-response pathway and biosorption capacity for conductive track formation in fungal mats, creating breadboard-like electronic substrates that could be compostable. This project integrates genetic engineering and thigmotropism to achieve custom-patterned metal deposition along the fungal mat. It expands synthetic biology by working with less common organisms such as N.crassa to robust fungal materials for electronics.
This project and similar existing proposals could help tackle the 62 million metric tons of annual e-waste, 80% of which is non-recyclable PCBs (Baldé et al., 2024). Biodegradable substrates could reduce landfill burden and explore more localized production. It mixes bioremediation, electronics and synthetic biology. Successful outcomes for MycoBoard would also further validate fungi as editable electronic chassis.
Even though MycoBoard involves genetic engineering for a beneficial and non-maleficient cause, copper or silver biosorption could mobilize metals if the fungal mats decompose improperly. Ethical considerations are key when working with living ecosystems and heavy metals. Possible containment methods for this could be auxotrophic markers, kill-switches, and lab-only strains that could prevent environmental release. Additionally to these, a key rule would be metal-loading protocols that use non-toxic concentrations. Throughout aim 2 and 3 future developments, iterative soil degradation assays are required to better understand copper toxicity thresholds and decomposition processes of the mats.
After presenting my research proposal during the Global Committed Listener presentation session, I received very useful feedback along with valuable references that helped me further understand MycoBoard’s positioning and technical framing (thank you to those who took the time to share these resources!). One particularly relevant publication by Rivnay et al. (2025) discusses how bioelectronics often requires high power and lacks the specificity and adaptability of some cells and tissues, and discussing that parallel advances in synthetic biology, biomaterials, and bioelectronics enable new opportunities in devices for regulated cell therapies, diagnostic tools, and next-generation robotics through biohybrid systems. Additionally, Lazaro-Vasquez and Vega (2019) demonstrated the use of mycelium composites with common digital fabrication techniques to replace plastic in electronics, specifically for inserting electronics in mycelium boards and making enclosures for electronics. However, their work focused only on enclosures and did not replace other components within the electronics; this is a gap that MycoBoard aims to address by engineering the fungal mat itself to form conductive tracks. On the other hand, some interesting advances are being done by The Rivnay Group’s work on organic bioelectronic materials that enable mixed ionic-electronic conduction for sensing and stimulation in biomedical settings (Rivnay Group, n.d.); and the Light Plate Apparatus (LPA), which could offer a potential platform for precisely controlling light-inducible expression of copper uptake and metallothionein genes in MycoBoard’s genetically engineered strains.
SECTION 4: EXPERIMENTAL DESIGN, TECHNIQUES, TOOLS, AND TECHNOLOGY
Detailed Plan for Aim 1 (Timeline: 2 weeks)
| Week | Day | Task |
|---|---|---|
| Week 1 | Day 1 | Retrieve N. crassa cmt CDS and Pccg-1 promoter from FungiDB; import to Benchling folder. Assemble linear cassette. Codon-optimize cmt for N. crassa. Annotate elements. Verify HA in-frame translation. |
| Day 2 | Split into Twist fragments. Export FASTA files. Upload fragments on Twist and revise correctedness of sequences. | |
| Day 3 | Fill out Excel and Google Forms for simulated Twist orders and Final Project proposals. | |
| Day 4 | Run NEB Cutter for restriction sites. Revise DNA sequences in relation to scientific literature resources. | |
| Day 5-6 | Literature revision for background and abstract sections of Final Project Documentation. | |
| Day 7 | Literature revision for metal absorption (silver vs. copper) - define best experimental scenario. | |
| Week 2 | Day 1-3 | Simulate copper-loading using experimental data from scientific literature and known Cu6 stoichiometry. |
| Day 3-4 | Validate benchmark for simulated copper-loading. | |
| Day 4-7 | Document restriction analysis, codon usage, and Twist order. Prepare transformation protocol outline. |
Techniques relevant to this project
- Lab Safety
- Bioethical Considerations
- DNA Construct Design
- Databases (e.g., GenBank, NCBI, Ensembl, and UCSC Genome Browser)
- Designing a Twist Order
- Use of Benchling
- Models and Notebooks
- Databases
Benchling DNA Construct Design: I used Benchling to assemble the linear cassette, by importing cmt CDS and Pccg-1 promoter from FungiDB, fusing C-terminal HA tag, and integrating hygR selection (see complete procedure here: https://pages.htgaa.org/2026a/sara-gaviria-escobar/projects/individual-final-project/index.html) . Annotation verified promoter-gene-terminator flow, codon optimization and translation viewer confirmed in-frame HA. The use of Benchling was key to validate the genetic logic for the cmt copper-responsive expression.
Twist Order Design:
The 4.5 kb Benchling cassette was longer than Twist’s single-fragment limit, so I split it into almost equal 2.4 kb and 2.1 kb pieces, keeping overhangs for future PCR or electroporation. This cost optimization avoided unnecessary backbone, targeting direct and easier spheroplast transformation for future development of the project.
How To Grow (Almost) Anything Industry Council companies associated with this final project
- Twist Biosciences/Ginkgo Bioworks (DNA synthesis)
- New England Biolabs (restriction enzyme analysis)
- Mycoworks (fungal materials precedent)
SECTION 5: Results & Quantitative Expectations
I chose to validate the DNA construct design for cmt overexpression in N. crassa by creating a complete Benchling cassette, checking reading frames and annotations, analyzing restriction sites, and deriving a literature-based copper-loading benchmark.
A detailed protocol for my validations:
- Obtained cmt (NCU05561) and Pccg-1 from FungiDB.
- Assembled Pccg-1=cmt-HA=hygR linear cassette.
- Codon-optimized cmt, verified HA fusion translation.
- Ran NEB Cutter to check there were no restriction enzyme sites. Results show no HA sites, 2 cmt sites (Eco0109I, MfeI), and multiple hygR sites.
- Simulated a Twist order ($359.92).
- Calculated Cu loading by researching experimental data in scientific literature.
The techniques for this protocol required:
- DNA Construct Design (Benchling assembly)
- Databases (FungiDB)
- Twist Order Design
- Models/Notebooks (Cu-loading calculation)
- Restriction Enzyme Analysis
The data obtained for validation was calculated based on existing scientific literature, and theoretical Cu biosorption values were calculated: Cu6 stoichiometry and 8% w/w value. Analysis also shows 54% dry mass as saturated cmt to match observed Cu, indicating broader biosorption contribution.
An unexpected challenge for this section was no direct cmt mg/cm² data found, which was resolved by using published stress-response values as upper bounds. The limitation, however, is that the estimate assumes uniform loading, which means future Western blots or similar methods are needed for cmt fraction validation.
To determine whether 0.8 mg of copper per cm² (measured from a fungal mat decomposing on soil according to scientific literature) falls below or above the toxicity threshold, the concentration must be converted to the standard unit of mg Cu per kg of dry soil. This calculation also assumes the fungal mat will decompose in the top 10 cm of soil (which is the typical active microbial zone) with a bulk density of approximately 1.3 g/cm³, 0.8 mg Cu/cm² over 10 cm depth equals a soil concentration of approximately 61.5 mg/kg (because 0.8 mg Cu ÷ 10 cm³ soil × 1.3 g/cm³ = 13 g soil = 0.013 kg; 0.8 / 0.013 ≈ 61.5 mg/kg). When compared to the cited thresholds, Shaw et al. (2020) observed loss of soil microbial functionality and community shifts above 200 mg Cu/kg, with no particular functional loss at or below 200 mg/kg. On the other hand, Rooney et al. (2006) reported plant toxicity EC50 values ranging from 36–536 mg/kg (in barley plants) and 22–851 mg/kg (in tomato plants) depending on soil properties, meaning effects typically emerge at concentrations higher than 60 mg/kg. Additionally, Caetano et al. (2016) derived soil screening values of 26.3–31.8 mg/kg for Cu based on multiple species and endpoints, no adverse effects are expected below these thresholds.
In conclusion, 61.5 mg/kg exceeds the preliminary screening value of ~30 mg/kg from Caetano et al. (2016) but is well below the 200 mg/kg functional threshold from Shaw et al. (2020) and the lowest plant EC50 values from Rooney et al. (2006). This means that while 0.8 mg/cm² is above the most conservative reference from my research, it remains under the threshold for significant microbial functionality loss (Shaw et al., 2020) and plant toxicity for most soils (Rooney et al., 2006). The fungal mat decomposition would not exceed the European Union’s agricultural warning limit of 200 mg/kg, but still needs further assessment depending on real copper concentrations from the actual mat grown at the lab.
SECTION 6: ADDITIONAL INFORMATION
References:
Baldé, C. P., Kuehr, R., Yamamoto, T., McDonald, R., D’Angelo, E., Althaf, S., … & Wagner, M. (2024). Global e-waste monitor 2024.
Beltramini, M., & Lerch, K. (1986). Primary structure and spectroscopic studies of Neurospora copper metallothionein. Environmental health perspectives, 65, 21.
Beltramini M, Lerch K. (1986). Primary structure and spectroscopic studies of Neurospora copper metallothionein. Environ Health Perspect. 65, 21-7. doi: 10.1289/ehp.866521. PMID: 3011391; PMCID: PMC1474700.
Carroll, A. M., Sweigard, J. A., & Valent, B. (1994). Improved vectors for selecting resistance to hygromycin. Fungal Genet. Newsl, 41(22), 135-143.
Caetano, A. L., Marques, C. R., Gonçalves, F., Da Silva, E. F., & Pereira, R. (2016). Copper toxicity in a natural reference soil: ecotoxicological data for the derivation of preliminary soil screening values. Ecotoxicology, 25(1), 163-177.
Cobbett, C., & Goldsbrough, P. (2002). Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Annual review of plant biology, 53(1), 159-182.
Danninger, D., Pruckner, R., Holzinger, L., Koeppe, R., & Kaltenbrunner, M. (2022). MycelioTronics: Fungal mycelium skin for sustainable electronics. Science Advances, 8(45), eadd7118.
Germann, U. A., & Lerch, K. (1987). Copper accumulation in the cell-wall-deficient slime variant of Neurospora crassa. Comparison with a wild-type strain. Biochemical journal, 245(2), 479-484.
Jabran M, Shafique MS, Abbas A, Han W, Ali MA, Gao L. (2025). Antifungal potential of silver and copper nanoparticles in suppressing Tilletia indica for Karnal bunt resistance in wheat. BMC Plant Biol. 25(1), 1517. doi: 10.1186/s12870-025-07547-x. PMID: 41199180; PMCID: PMC12590600.
Kumar, K. S., Dayananda, S., & Subramanyam, C. (2005). Copper alone, but not oxidative stress, induces copper–metallothionein gene in Neurospora crassa. FEMS microbiology letters, 242(1), 45-50.
Lazaro-Vasquez, E. S., & Vega, K. (2019). From plastic to biomaterials: Prototyping DIY electronics with mycelium. In Adjunct Proceedings of the 2019 ACM International Joint Conference on Pervasive and Ubiquitous Computing and Proceedings of the 2019 ACM International Symposium on Wearable Computers (UbiComp/ISWC ‘19 Adjunct) (pp. 308–311). ACM. https://doi.org/10.1145/3341162.3343808
Lerch, K. (1980). Copper metallothionein, a copper-binding protein from Neurospora crassa. Nature, 284(5754), 368-370.
Mohan, C. R., Kandasamy, R., & Kabiriyel, J. (2024). Correlation between electrical conductivity and antibacterial activity of chitosan-stabilized copper and silver nanoparticles. Carbohydrate Polymer Technologies and Applications, 7, 100503.
Nargang, C. E., & Nargang, F. E. (1996). Procedure for preparing and transforming spheroplasts of Neurospora crassa.
Nielson KB, Atkin CL, Winge DR. (1985). Distinct metal-binding configurations in metallothionein. J Biol Chem. 260(9), 5342-50. PMID: 3988757.
Ohrnberger, J., & Akins, R. A. (1995). Cloning of the copper-inducible metallothionein (cmt) promoter from Neurospora crassa. Fungal Genetics Reports, 42(1), 57-58.
Pazirandeh, M., Chrisey, L.A., Mauro, J.M. et al. (1995). Expression of the Neurospora crassa metallothionein gene in Escherichia coli and its effect on heavy-metal uptake. Appl Microbiol Biotechnol 43, 1112–1117. https://doi.org/10.1007/BF00166934
Ploessl, D. (2022). Developing nuclear and mitochondrial DNA editing techniques for engineering yeasts as novel microbial factories and disease models (Doctoral dissertation, Iowa State University).
Rai, M., Bonde, S., Golinska, P., Trzcińska-Wencel, J., Gade, A., Abd-Elsalam, K. A., … & Ingle, A. P. (2021). Fusarium as a novel fungus for the synthesis of nanoparticles: mechanism and applications. Journal of Fungi, 7(2), 139.
Riquelme, M., Aguirre, J., Bartnicki-García, S., Braus, G. H., Feldbrügge, M., Fleig, U., … & Fischer, R. (2018). Fungal morphogenesis, from the polarized growth of hyphae to complex reproduction and infection structures. Microbiology and Molecular Biology Reviews, 82(2), 10-1128.
Rivnay, J., Raman, R., Robinson, J. T., & et al. (2025). Integrating bioelectronics with cell-based synthetic biology. Nature Reviews Bioengineering, 3, 317–332. https://doi.org/10.1038/s44222-024-00262-6
Rivnay Group. (n.d.). Organic bioelectronic materials, devices and systems. Northwestern University. Retrieved May 23, 2026, from https://rivnay.northwestern.edu/
Robinson, J., Munagala, S. P., Arjunan, A., Simpson, N., Jones, R., Baroutaji, A., … & Lyall, I. (2022). Electrical conductivity of additively manufactured copper and silver for electrical winding applications. Materials, 15(21), 7563.
Romeyer, F. M., Jacobs, F. A., & Brousseau, R. (1990). Expression of a Neurospora crassa metallothionein and its variants in Escherichia coli. Applied and environmental microbiology, 56(9), 2748-2754.
Rooney, C. P., Zhao, Fangjie, McGrath, Steve (2006) Soil factors controlling the expression of copper toxicity to plants in a wide range of European soils. Environmental Toxicology And Chemistry, 25 (3). pp. 726-732. ISSN 0730-7268
Singh, K., Sharma, S., Kalia, A., & Manchanda, P. (2026). Advancement in Mushroom Transformation: From Conventional Techniques to Modern Genetic Engineering. Journal of Basic Microbiology, 66(1), e70132.
Shaw, J. L. A., Ernakovich, J. G., Judy, J. D., Farrell, M., Whatmuff, M., & Kirby, J. (2020). Long-term effects of copper exposure to agricultural soil function and microbial community structure at a controlled and experimental field site. Environmental Pollution, 263, 114411.
Staben, C., Jensen, B., Singer, M., Pollock, J., Schechtman, M., Kinsey, J., & Selker, E. (1989). Use of a bacterial hygromycin B resistance gene as a dominant selectable marker in Neurospora crassa transformation. Fungal Genetics Reports, 36(1), 79.
Subramanyam, C., Venkateswerlu, G., & Rao, S. L. N. (1983). Cell wall composition of Neurospora crassa under conditions of copper toxicity. Applied and environmental microbiology, 46(3), 585-590.
Suresh, K., & Subramanyam, C. (1998). Polyphenols are involved in copper binding to cell walls of Neurospora crassa. Journal of inorganic biochemistry, 69(4), 209-215.
Tabor Lab. (n.d.). Light Plate Apparatus (LPA): Open-source instrument for optogenetics. Rice University. Retrieved May 23, 2026, from https://taborlab.github.io/LPA-hardware/index.html
Varshney, R., Bhadauria, S., & Gaur, M. S. (2012). A review: biological synthesis of silver and copper nanoparticles. Nano Biomedicine & Engineering, 4(2).
Wang, Z., Bartholomai, B. M., Loros, J. J., & Dunlap, J. C. (2023). Optimized fluorescent proteins for 4-color and photoconvertible live-cell imaging in Neurospora crassa. Fungal Genetics and Biology, 164, 103763.
Yoon, K. Y., Byeon, J. H., Park, J. H., & Hwang, J. (2007). Susceptibility constants of Escherichia coli and Bacillus subtilis to silver and copper nanoparticles. Science of the Total Environment, 373(1), 572–575.
Supply list:
| Item | Cost ($) |
|---|---|
| Twist DNA synthesis | 360 |
| N. crassa spores | 100 |
| Hygromycin B | 50 |
| Vogel’s medium + CuSO4 | 30 |
| Laser-cut/3D printed acrylic molds | 50 |
| Multimeter + LED/resistor | 20 |
TOTAL: approx. 610 $