Week 7 HW: Genetic Circuits Part 2
Part 1: Intracellular Artificial Neural Networks (IANNs)
Advantages of IANNs over Boolean genetic circuits
IANNs offer three advantages over Boolean genetic circuits.
- Graded signaling. They operate on continuous intracellular signals rather than discrete ON/OFF states, enabling weighted summation, nonlinear activation, and universal function approximation. Weiss-coauthored neuromorphic circuits demonstrated these capabilities through analog computation, soft majority voting, and ternary switching in living cells.
- Tunable decision boundaries. Effective weights and biases can be adjusted by modifying stoichiometry, promoter strength, or recognition-site placement; no topological redesign is required. The PERSIST endoRNase system illustrates this: the same RNase acts as a repressor or activator depending on whether the target site is in the 5′ UTR or 3′ UTR.
- Greater expressive power. Multilayer IANNs can represent smooth classifiers and nonlinearly separable response surfaces that Boolean truth tables cannot efficiently encode.
A useful application
An autonomous cell-state classifier for stem-cell differentiation would be a strong use case. The IANN would integrate sensors for an endothelial-intermediate RNA signature ($x_1$), residual pluripotency ($x_2$), and off-target lineage markers ($x_3$), computing a weighted sum
$$z = w_1 x_1 - w_2 x_2 - w_3 x_3 + b$$
passed through a nonlinear output node that drives a fluorescent reporter or a differentiation factor. Weiss and colleagues used endoRNase-mediated miRNA sensors in a similar fashion to monitor cell-state transitions and guide multistep hiPSC differentiation toward a hematopoietic lineage.
Limitations:
- Resource loading in mammalian cells (Weiss 2020 showed competing modules can reduce unregulated gene expression by up to 70%).
- RNase saturation and cross-cleavage at high enzyme ratios, as observed in PERSIST cascades.
- Stochastic weight variation across cells from poly-transfection.
- The 650 ng total-DNA constraint imposed by the class protocol, which the supplied two-layer design already saturates.
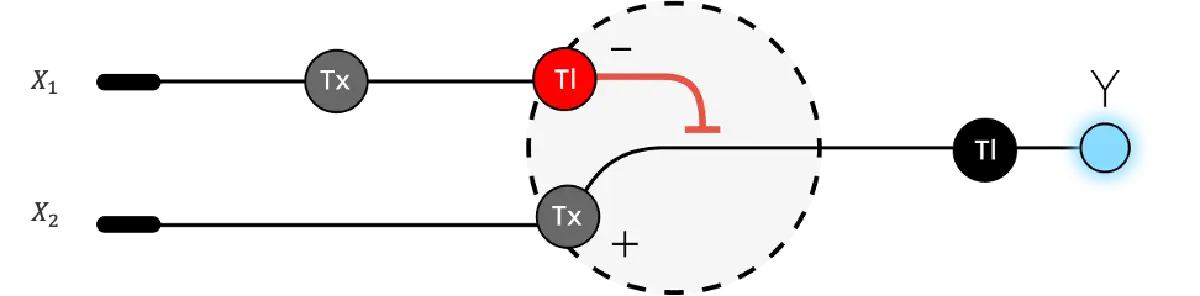
Single-layer IANN diagram
The diagram below shows an intracellular single-layer perceptron where the X1 input is DNA encoding the Csy4 endoribonuclease and the X2 input is DNA encoding a fluorescent protein output whose mRNA is regulated by Csy4. (Tx: transcription; Tl: translation.)
Multilayer IANN diagram
The diagram below shows a two-layer intracellular perceptron built from the supplied parts.
| Layer | Function |
|---|---|
| Layer 1 | Input DNA X1 encodes Csy4. Csy4 protein cleaves the Csy4 recognition site on the hidden-layer transcript (Csy4_rec_CasE), repressing it and producing the hidden-node output H = CasE. |
| Layer 2 | CasE protein acts on the CasE recognition site in the output transcript (CasE_rec_mNeonGreen), repressing it to produce the fluorescent output Y = mNeonGreen. |
Both RNase links are drawn as repression to match the supplied single-layer example. In PERSIST-style designs, the sign of each edge can be inverted by repositioning the recognition site from a 5′-UTR OFF configuration to a 3′-UTR ON configuration.
In the provided spreadsheet, this design corresponds to:
- X1 =
Csy4 + mKO2 - X2 =
Csy4_rec_CasE + eBFP2 - Bias =
CasE_rec_mNeonGreen
It consumes the full 650 ng class DNA limit.

Part 2: Fungal Materials
Examples of existing fungal materials
Most fungal materials are mycelium-based.
| Product | Use | Notes |
|---|---|---|
| Mycelium packaging (Ecovative, used by Dell) | Styrofoam replacement | Made by inoculating agricultural waste with fungal spores and molding it into custom shapes. |
| Mycelium leather (MycoWorks, Bolt Threads Mylo) | Leather replacement | Roughly 70% less water and 68% lower greenhouse gas emissions than cattle leather. |
| Construction and insulation panels | Building materials | Fire-resistant, with favorable thermal conductivity and sound absorption. |
| Compostable foams (Ecovative) | Packaging, padding | |
| Fungal protein foods (Mycorena) | Food |
Common advantages over traditional counterparts: biodegradability, use of waste feedstocks, reduced environmental impact.
Common disadvantages: limited mechanical performance (mycelium compressive strength is around 0.1 to 0.2 MPa vs. 17 to 28 MPa for concrete), moisture susceptibility, batch-to-batch variability, scaling difficulties, and (in the case of leather substitutes) cost issues that have forced some manufacturers to shut down.
Why genetically engineer fungi?
Engineering targets in fungi include:
- Modifying cell-wall biosynthetic genes (chitin synthase, alpha-glucan synthase, acetyltransferases) to tune material properties at the genome level.
- Activating silent secondary-metabolite gene clusters via synthetic transcription factors or heterologous expression in hosts like Aspergillus oryzae.
- Producing non-native compounds (cannabinoids, biofuels, therapeutic proteins).
- Embedding synthetic gene circuits into mycelium to create stimulus-responsive living materials.
Advantages over bacteria for synthetic biology
- Eukaryotic post-translational machinery. Glycosylation, disulfide bonds, and proteolytic processing are needed for functional human therapeutic proteins.
- High secretion capacity. Filamentous fungi secrete 10 to 1,000× more protein than bacterial hosts.
- Native handling of complex gene clusters. Fungi harbor secondary-metabolite pathways with large intron-containing gene clusters that bacterial systems cannot properly express.
- 3D growth. Mycelium grows into three-dimensional networks usable directly as structural materials. No bacterial system offers this.
- Lignocellulosic feedstocks. Fungi thrive on waste streams that most bacteria cannot degrade.
Tradeoffs. Slower growth, less well-characterized genetics, and a synthetic biology toolkit that remains less mature than what is available for E. coli. Recent efforts like the Fungal Modular Cloning Toolkit (96 standardized parts for filamentous fungi) are narrowing this gap.
Part 3: First DNA Twist Order
All draft Round 0 constructs are deposited in Benchling at https://benchling.com/seanmurp/f_/KopGo3fSDI-htgaa_final_project/, organized into sub-folders for Round 0 constructs (by reporter: sfGFP, mCherry, NanoLuc) and controls.
The library comprises 80 unique 20-nt T7 promoter-spacer variants (positions +1 to +20), each paired with three reporters (sfGFP, mCherry, NanoLuc), yielding 240 test constructs plus 9 controls (dead-promoter negatives, no-RBS negatives, and synonymous codon-variant sfGFP controls).
Spacers are drawn from five design categories:
- Published reference variants (WT T7, T7Max, T7c62, T7#4).
- Systematic ITS mutagenesis at positions +1 to +6.
- RBS / translation-efficiency variants.
- Context-interaction variants designed to produce reporter-dependent expression differences.
- Random space-filling variants for unbiased landscape coverage.
Construct architecture
All constructs are designed as linear DNA fragments:
Total lengths range from ~720 bp (NanoLuc) to ~920 bp (sfGFP). The 59-bp flanking buffers protect against residual RecBCD exonuclease activity in the BL21 Star lysate, per Ginkgo’s recommendation of 50 to 80 bp padding for linear DNA templates in their cell-free system. Constructs will be synthesized as linear gene fragments (e.g., via Twist Bioscience) and used directly as CFPS templates at 15 to 20 nM, with no plasmid cloning.