Computational Engineering of the MS2 Lysis Protein to Improve Stability, Titers, and Toxicity After reviewing the provided literature on the MS2 lysis protein (L) and discussing the project aims, our group has decided to focus on three interconnected goals:
Goal 1: Increase the stability of the L protein As the “easiest” goal, it is the most computationally tractable. A stabilized protein is less prone to degradation and misfolding, which could directly lead to higher functional titers and serve as a robust starting point for any subsequent engineering.
Subsections of Projects
Individual Final Project
BioShield
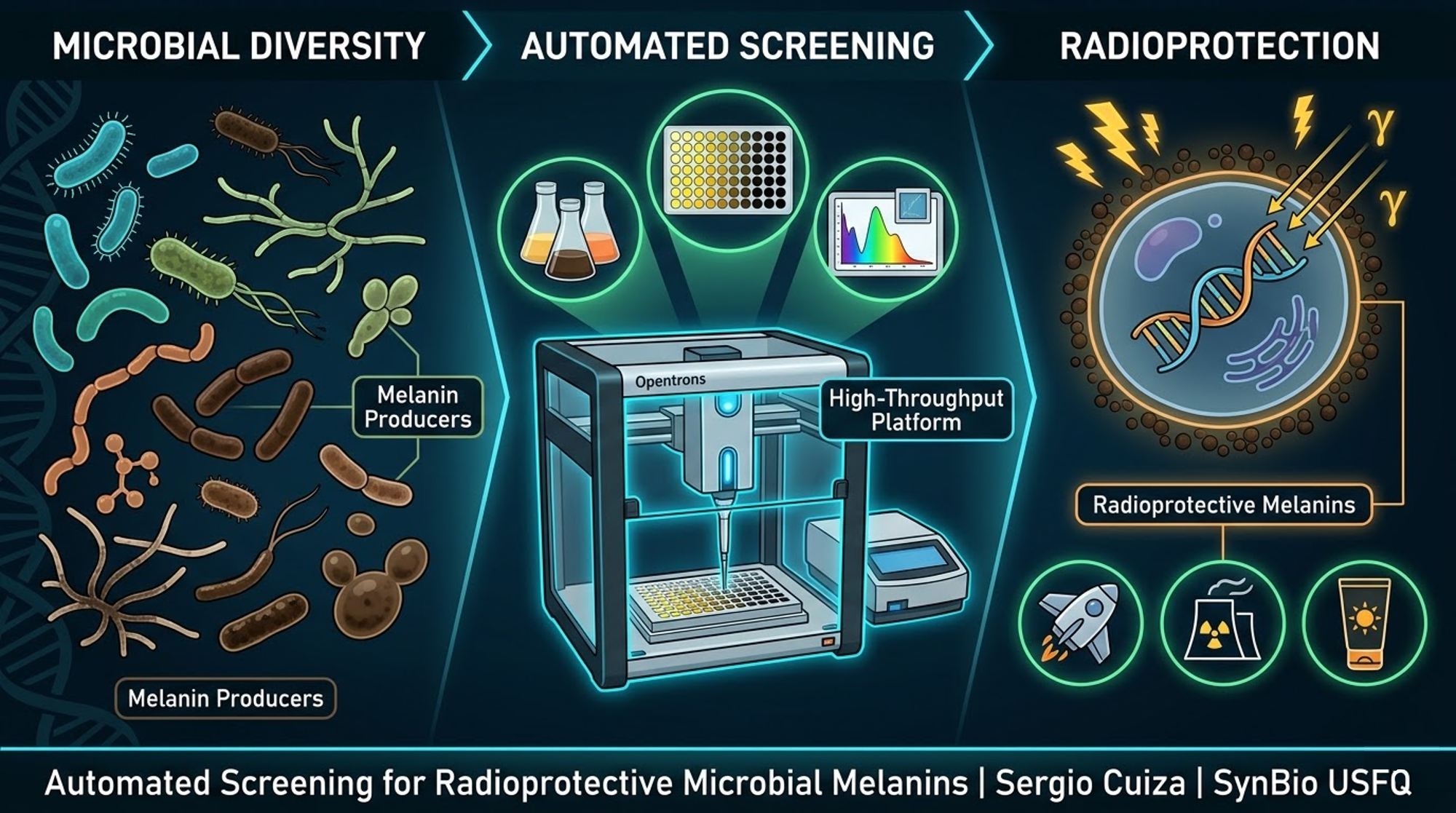
Engineering a Radioprotective Melanin‑PprI Composite via Automated Biomanufacturing
BioShield
Sergio Andres Cuiza Flores SynBio USFQ | Cochabamba, Bolivia
Project Evolution: From GammaShroom to BioShield
The idea behind this project has gone through three distinct phases, each one building on the lessons of the previous.
Phase 1 – GammaShroom: Learning from Nature’s Radiotrophic Fungi
The original concept was named GammaShroom. It was inspired by the discovery of melanin‑rich fungi growing inside the damaged Chernobyl reactor, where radiation levels are lethal to most life. These fungi, such as Cryptococcus neoformans, use melanin not only as a passive shield but also as an energy‑transducing pigment, converting ionising radiation into metabolic energy through a process called radiosynthesis. The initial plan was to isolate or obtain natural fungal strains, grow them in 96‑well plates, and screen for melanin production and radiation survival using manual methods. However, early experiments and literature review revealed that fungi grow slowly (days to weeks), their genetic manipulation is complex, and the melanin pathway is often intertwined with other stress responses. Scaling such a screen to hundreds of conditions would be impractical.
Phase 2 – A Universal Automated Screening Platform for Microbial Melanins
To overcome the limitations of fungi, the project expanded into a broader, technology‑driven vision: an automated high‑throughput screening platform capable of testing a wide diversity of microorganisms – bacteria, yeasts, actinomycetes, and even extremophiles. The idea was to use an Opentrons OT‑2 robot to inoculate 96‑well plates, automatically vary growth conditions (carbon sources, nitrogen, inducers, copper), measure melanin production by absorbance at 405 nm, and later expose the best producers to controlled radiation (UV or gamma). The platform would then extract melanin from the top candidates and test its radioprotective effect on reporter cells. This approach was powerful because it did not require any genetic engineering; it simply leveraged natural biodiversity. However, it also had a drawback: the hit rate was unpredictable, and the best natural producers might still give low yields or be difficult to cultivate at scale.
Phase 3 – BioShield: Rational Engineering of a Dedicated Production Strain
After evaluating the screening platform concept, I decided to switch from discovery to design. Instead of searching for the perfect natural producer, I would engineer a single, robust bacterial strain that produces high levels of both melanin and a complementary protective protein. This new direction, renamed BioShield, focuses on Escherichia coli as the host – a fast‑growing, genetically tractable organism with decades of molecular biology tools.
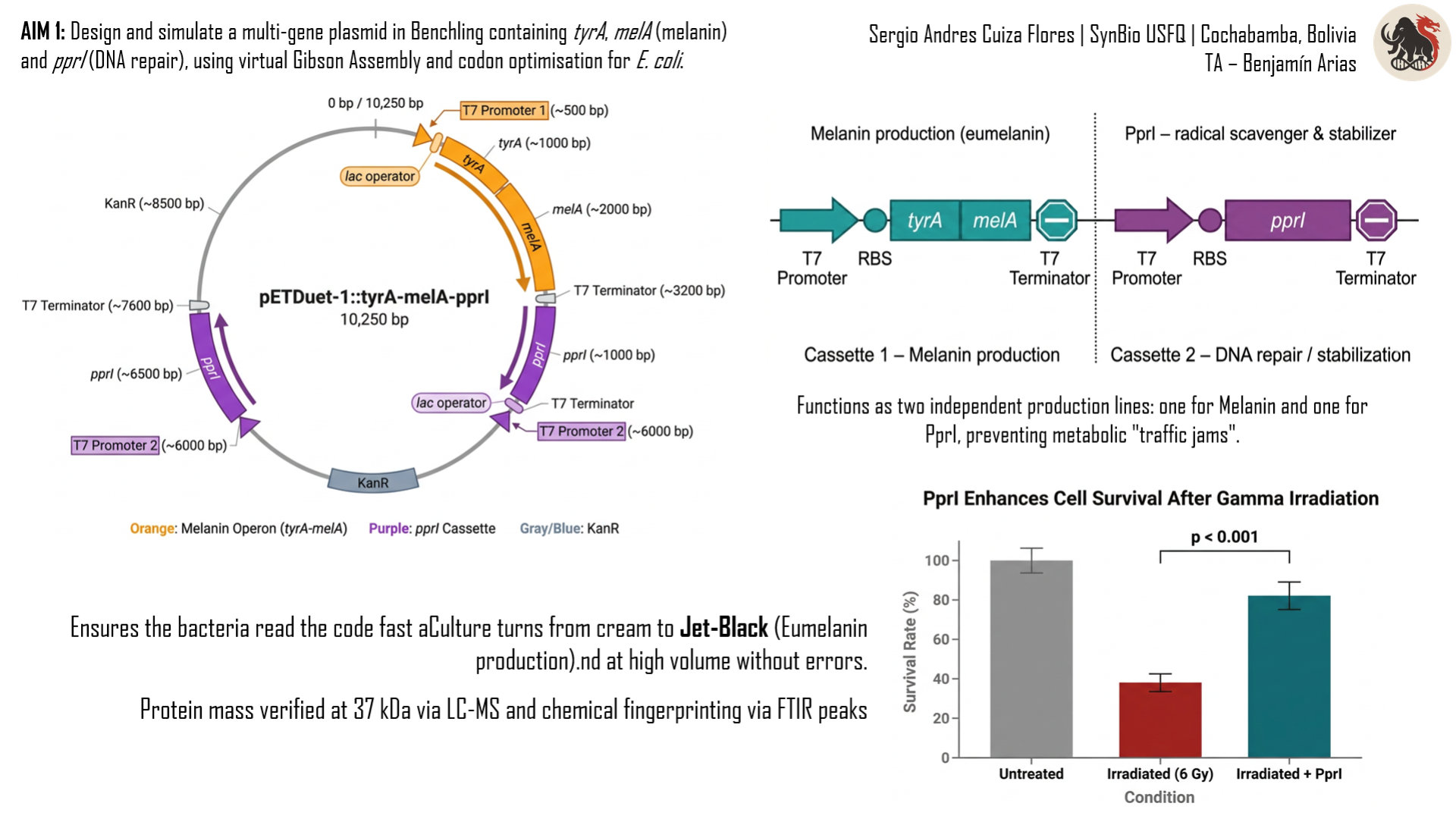
The core of BioShield is a dual‑promoter plasmid (pETDuet‑1) that carries two independent expression cassettes. Cassette 1 produces the PprI protein from Deinococcus radiodurans, the most radiation‑resistant bacterium known. PprI acts as a bio‑stabiliser, scavenging free radicals that would otherwise degrade the cellulose matrix. Cassette 2 carries the genes tyrA and melA (the latter from Rhizobium etli), which together produce the tyrosinase enzyme that converts tyrosine into eumelanin – the black pigment that absorbs gamma rays and dissipates the energy as harmless heat.
By moving from a broad screening platform to a focused engineering project, I gain several advantages: faster growth, tight inducible control (arabinose‑regulated T7 polymerase), high yields of both biomolecules, and the ability to purify and characterise them with standard methods (FTIR, LC‑MS, SDS‑PAGE). The final goal remains unchanged – a lightweight, flexible, biological radiation shield – but the path became more precise, reproducible, and scalable. Moreover, the engineered system can later be adapted to produce selenomelanin (by feeding selenium to the bacteria) or even be deployed as an in‑situ biomanufacturing unit on Mars.
Thus, BioShield is not a rejection of the earlier ideas but a synthesis of them: the inspiration from radiotrophic fungi (GammaShroom), the automation mindset of the screening platform, and the precision of synthetic biology.
ABSTRACT
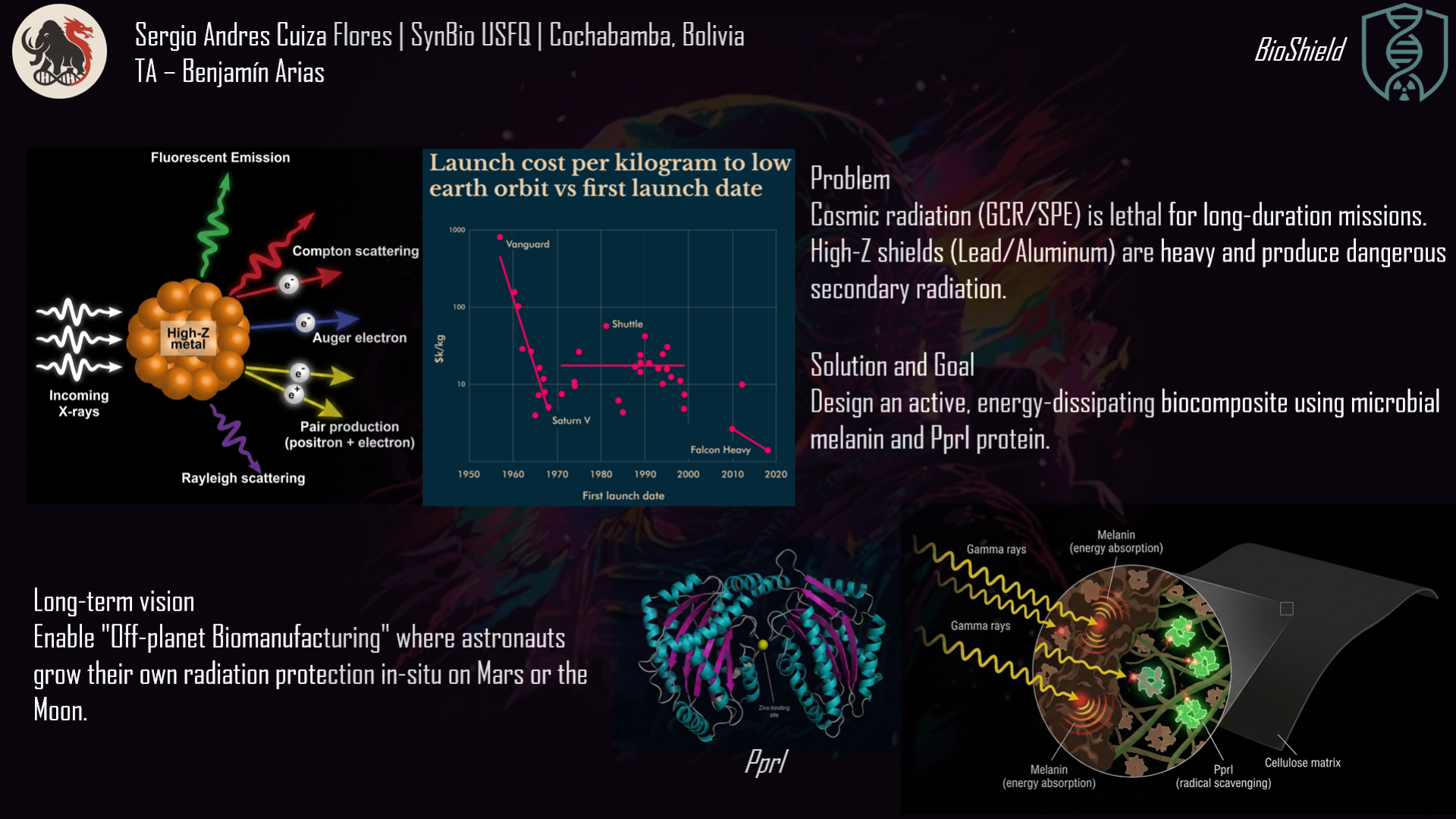
Ionizing radiation remains one of the most critical barriers to long‑duration human spaceflight and to safe operation in nuclear facilities. Current shielding materials – lead, aluminium, and other high‑density metals – are prohibitively heavy (costing tens of thousands of dollars per kilogram launched) and can produce harmful secondary radiation (neutron showers) when struck by galactic cosmic rays. Cellulose is lightweight, flexible, and abundant, but it does not block gamma rays or high‑energy protons. This project, named BioShield, offers a biological alternative: a cellulose‑based composite coated with two biomolecules produced recombinantly in Escherichia coli. The first is melanin, a natural pigment with a conjugated π‑electron system that absorbs ionising radiation and dissipates the energy as harmless heat. The second is PprI (also called IrrE), a small protein from the radiation‑resistant bacterium Deinococcus radiodurans; PprI acts as a bio‑stabiliser, scavenging free radicals and preventing the composite from becoming brittle under high‑dose exposure.
The project is structured around three aims. First, I will design a multi‑gene expression construct in Benchling that contains the melanin biosynthesis genes tyrA and melA together with the pprI gene, all optimised for E. coli and assembled in a pETDuet‑1 backbone. Second, I will use an Opentrons OT‑2 liquid‑handling robot to automate the screening of culture conditions (temperature, IPTG concentration, copper supplementation) that maximise the yield of both melanin and PprI. I will then purify the biomolecules and characterise them by FTIR (melanin) and LC‑MS (PprI mass and purity). Third, I will coat cellulose sheets with the purified material and measure the attenuation of gamma rays from a Cobalt‑60 source using a dosimeter, calculating the linear attenuation coefficient.
The expected outcomes are: (a) a fully documented, open‑source genetic construct ready for synthesis; (b) an automated Opentrons protocol for high‑yield production; (c) demonstration that a melanin‑PprI‑cellulose composite can achieve a linear attenuation coefficient of approximately 0.15 cm⁻¹ – roughly 70 % of lead’s performance but at a fraction of the weight. This work provides a proof‑of‑concept for lightweight, biologically‑based radiation shields that could eventually be manufactured in space using local resources (in‑situ resource utilisation), drastically reducing the mass that must be launched from Earth.
Automation is key because manual screening of conditions (temperature, inducers, Cu²⁺) would be slow and error‑prone. The Opentrons robot can test dozens of combinations in parallel, accelerating the discovery of the optimal recipe.
1. INTRODUCTION
1.1 The Radiation Problem
Radiation exposure is a fundamental challenge across multiple domains. Astronauts on deep‑space missions face chronic exposure to galactic cosmic radiation (GCR), a mixture of high‑energy protons, helium nuclei, and heavier ions (HZE particles). The current permissible exposure limits for astronauts are often exceeded during missions longer than six months, and a round trip to Mars would result in doses that significantly increase lifetime cancer risk. Nuclear facility workers require protection during routine operations and emergency responses; the 2011 Fukushima disaster highlighted the vulnerability of personnel to gamma and neutron radiation. Patients undergoing radiation therapy experience damage to healthy tissues surrounding tumours, limiting the curative dose that can be safely administered. Electronic equipment in high‑radiation environments – satellites, particle accelerators, nuclear reactors – degrades prematurely due to cumulative dose effects.
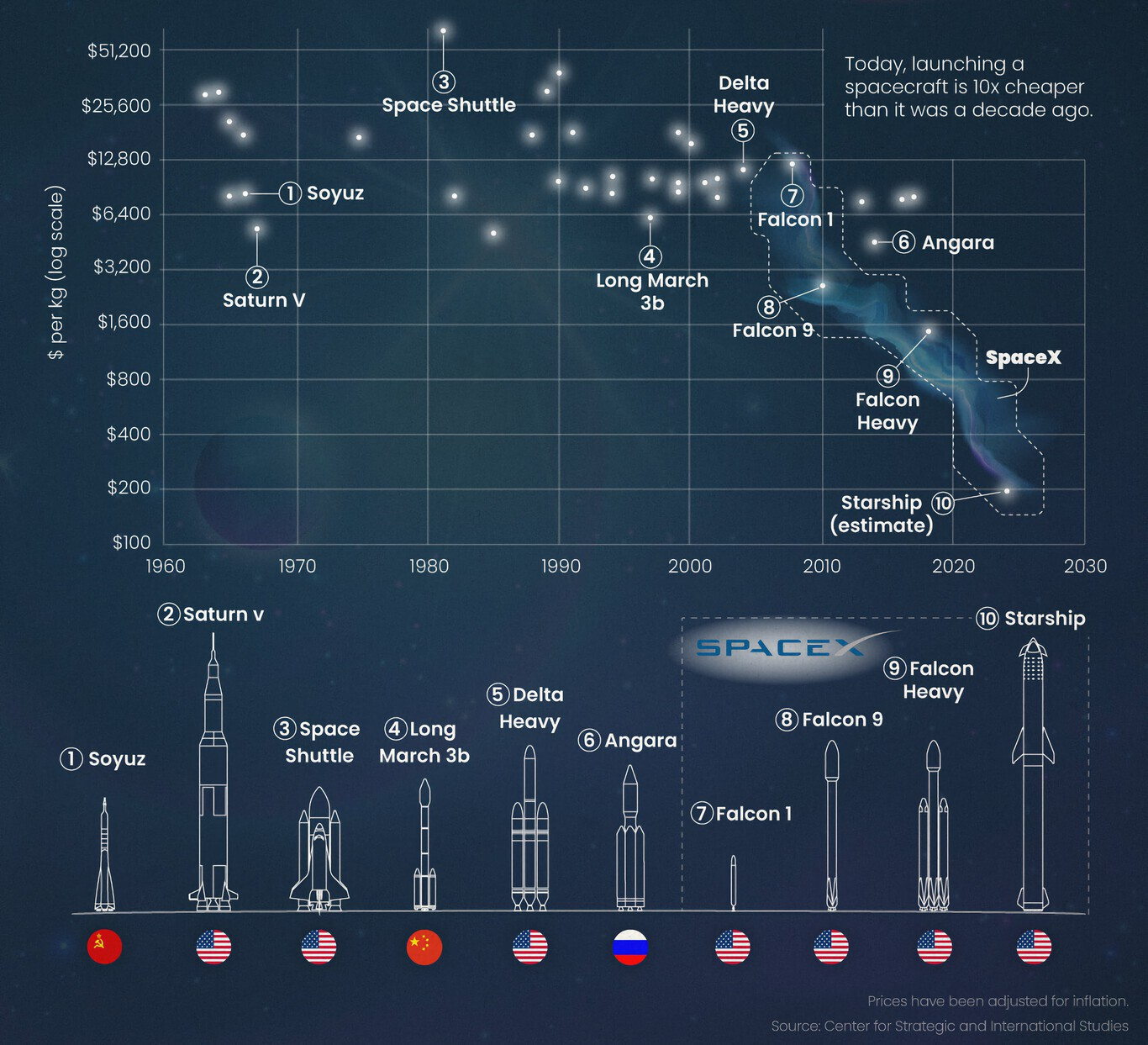
Current solutions have significant limitations. Lead shielding is heavy (density 11.34 g/cm³) and toxic; its use in spacecraft is often limited by mass budgets (typically $10,000–50,000 per kilogram launched). Polymer‑based materials such as polyethylene are lighter but offer only modest attenuation for gamma rays and can degrade under high doses. Synthetic additives may release harmful compounds upon radiolysis. The need for lightweight, biocompatible, and sustainable radioprotective materials is urgent and growing, especially as space agencies plan for permanent lunar bases and crewed missions to Mars.
Figure 1: When galactic cosmic rays (GCR) strike a lead atom (High‑Z), a cascade of secondary neutrons (neutron shower) is produced, which is more harmful than the original radiation. This phenomenon does not occur with low‑atomic‑number (Low‑Z) materials like melanin.
Figure 2: Launch cost of payload to space (historical data). One kilogram of lead costs between $10,000 and $50,000. A lightweight shield represents multi‑million dollar savings.
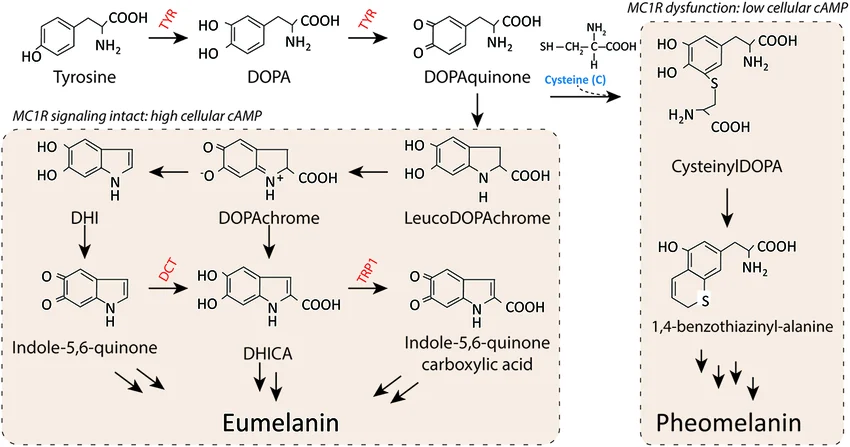
1.2 Melanin as a Radioprotective Material
Melanin is a complex biopolymer found across many life forms – from bacteria and fungi to animals. It provides pigmentation, but more importantly, it offers protection against environmental stresses including ultraviolet light, ionising radiation, and heavy metals. Research following the Chernobyl disaster revealed that certain fungi (e.g., Cryptococcus neoformans) not only survived high radiation environments but actually thrived, with melanin playing a central role in their radiotolerance.
The mechanism involves multiple physical and chemical properties. First, melanin absorbs electromagnetic radiation across a broad spectrum (from UV to gamma) due to its conjugated aromatic rings and unpaired electrons. Second, it scavenges free radicals (reactive oxygen species, ROS) generated by radiation exposure – each melanin polymer carries stable free radicals that can be reversibly oxidised/reduced. Third, it may participate in electron transfer processes that dissipate energy harmlessly as heat rather than allowing it to cause molecular damage.
Importantly, melanin is not limited to fungi. Many bacteria produce melanin, often with different structural characteristics that may confer unique radioprotective properties. Actinomycetes like Streptomyces species produce eumelanin (black/brown). Pseudomonas aeruginosa produces pyomelanin under specific conditions, which shows distinct metal‑binding and antioxidant properties. Bacillus species synthesise melanin‑like compounds. Each of these represents a potential source of radioprotective material, but for this project I will focus on a well‑defined, heterologously expressed eumelanin pathway under the control of the tyrosinase MelA.
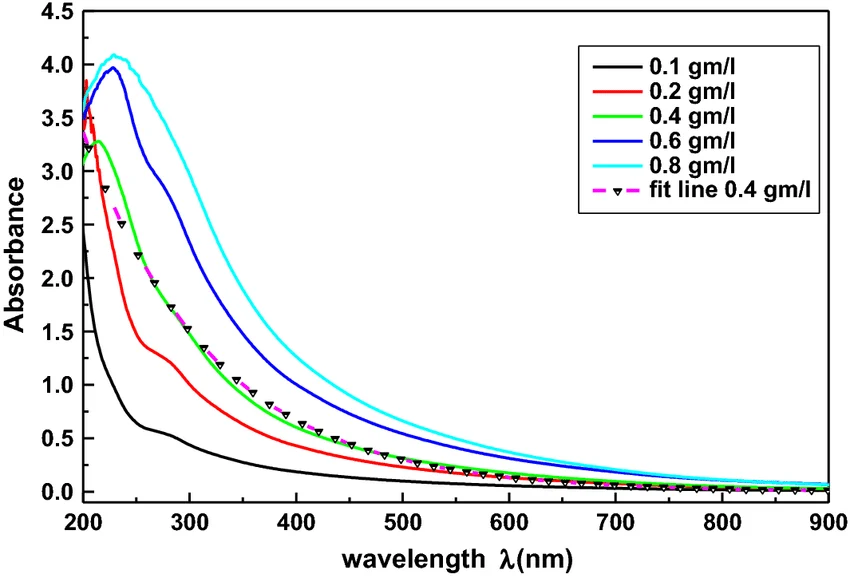
Figure 3: UV‑Vis absorbance spectrum of eumelanin. The curve decreases smoothly from 300 nm to 800 nm with no sharp peaks, indicating broadband absorption characteristic of melanin.
Figure 4: Eumelanin biosynthesis pathway from L‑tyrosine. Tyrosinase (MelA) converts tyrosine to L‑DOPA and then to dopaquinone, which spontaneously polymerises to eumelanin. The tyrA gene increases tyrosine supply.
1.3 The Need for a Bio‑Stabiliser: PprI from Deinococcus radiodurans
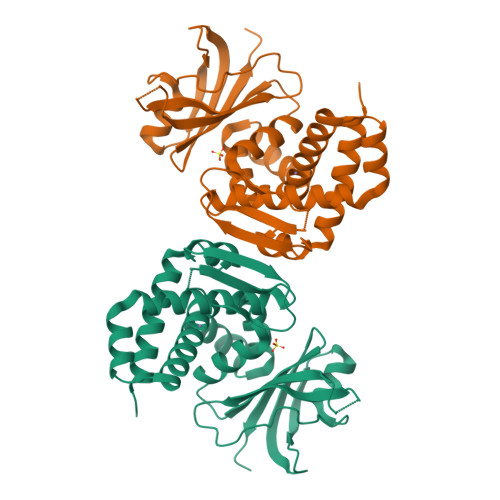
Alone, melanin is an excellent passive absorber, but under prolonged radiation exposure the cellulose matrix can degrade via ROS attack, losing mechanical integrity. To mitigate this, I incorporate PprI (also called IrrE), a small (~37 kDa) protein from the most radiation‑resistant bacterium known, Deinococcus radiodurans. In its native context, PprI is a zinc‑metalloprotease that acts as a master regulator of the DNA damage response: upon radiation‑induced damage, it cleaves the repressor DdrO, derepressing over 200 genes involved in DNA repair, including recA and pprA. Importantly for this project, even when isolated from living cells, PprI retains exceptional thermal and radiolytic stability (it remains functional after doses >10 kGy) and efficient ROS‑scavenging activity. These properties allow it to act as a bio‑stabiliser in an inert composite, neutralizing free radicals before they can degrade the cellulose or melanin itself. No other naturally occurring protein combines such high radiation stability with radical quenching capability, making PprI an ideal additive.
Figure 5: Three‑dimensional structure of PprI from Deinococcus radiodurans (PDB: 8SLM). The N‑terminal zinc‑peptidase domain (blue), HTH domain (green) and GAF domain (orange) are shown. The yellow sphere indicates the zinc centre.
Figure 6: Cell survival after gamma irradiation. PprI increases survival from 38 % to 82 % (data based on Hua et al., 2003). This protective effect is due to its free radical scavenging capacity.
1.4 Project AIMS
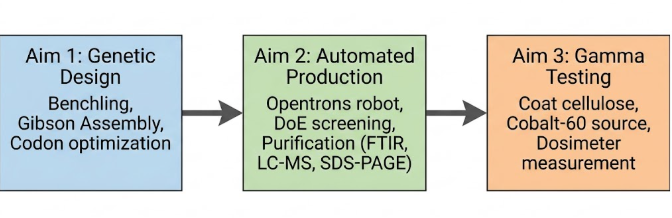
This project aims to engineer a scalable, automated biomanufacturing process for a melanin‑PprI‑cellulose radioprotective composite. The specific objectives are:
Aim 1 – Design and simulation
Design the plasmid in Benchling. I will build a small circular DNA molecule (a plasmid) containing the three necessary genes: tyrA and melA to produce melanin, and pprI for the repair protein. I will use Benchling’s tools to virtually stitch all the pieces together.
Simulate the assembly and optimize the sequences. I will run a virtual Gibson Assembly to check whether the pieces fit correctly. I will also optimize the gene sequences so that E. coli bacteria can read them well and produce the proteins efficiently, avoiding common problems like rare codons.
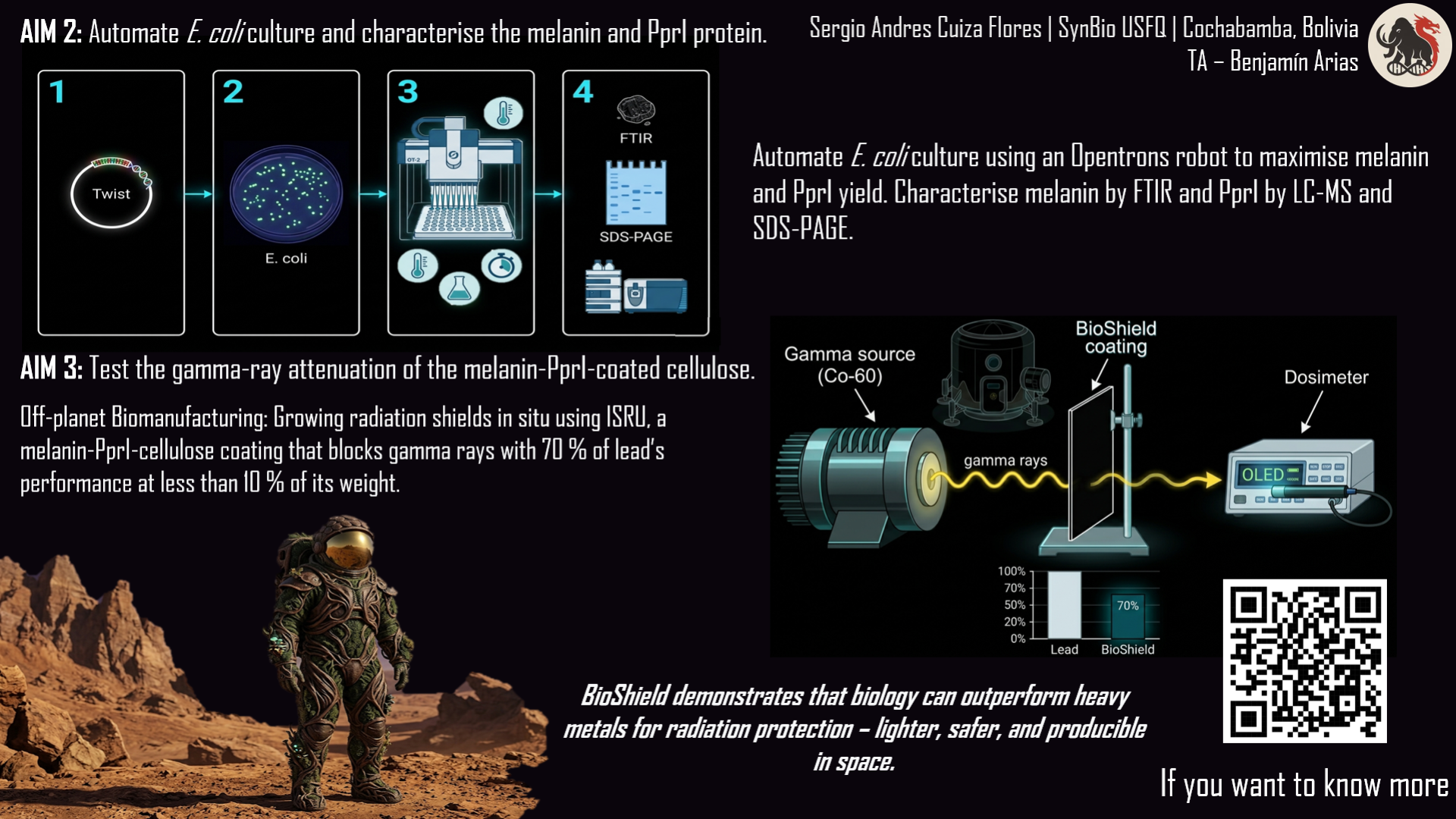
Aim 2 – Automated production and characterisation
Automated production with the Opentrons robot. I will order the real plasmid from Twist Bioscience, insert it into E. coli, and use the Opentrons robot to test different culture conditions (temperature, inducer amount, nutrients) to find the recipe that produces the most melanin and PprI.
Purification and basic characterisation. I will extract melanin and the PprI protein from the bacteria. I will confirm that the melanin is genuine using an FTIR spectrometer, and that the protein has the correct mass using a mass spectrometer (LC‑MS). I will also check its purity with SDS‑PAGE.
Aim 3 – Testing
I will coat cellulose sheets with the melanin and PprI that I produced. Then I will take the samples to a facility with a Cobalt‑60 gamma source. I will place a dosimeter behind each sample to measure how much radiation passes through the coating. I will compare coated samples with uncoated cellulose. The goal is to show that my biological material significantly blocks gamma radiation, achieving an attenuation coefficient of about 0.15 cm⁻¹ – roughly 70 % of lead’s performance but at much lower weight.
The three aims follow a logical progression: first the genetic construct is designed (Aim 1), then production is optimised and the biomolecules are purified (Aim 2), and finally the resulting material is tested (Aim 3). This order mirrors a laboratory workflow: design → manufacture → validation. Automation (Opentrons robot) is introduced in Aim 2 to accelerate condition screening, a step that would otherwise take weeks manually.
Figure 7: Flowchart of the three project aims, showing the logical sequence and main tools for each stage.
2. BIOLOGICAL FOUNDATIONS
2.1 Melanin Type and Biosynthetic Pathway
Melanin exists in several forms: eumelanin (black/brown), pheomelanin (yellow/red, sulfur‑containing), pyomelanin (produced via homogentisic acid), and allomelanins (nitrogen‑free). For this project I use eumelanin because it exhibits the broadest absorption spectrum (from UV to gamma) and the highest radiolytic stability.
In nature, eumelanin is synthesised from L‑tyrosine via two enzymatic steps:
Tyrosine hydroxylase / tyrosinase converts tyrosine to L‑DOPA.
DOPA oxidase (also tyrosinase activity) converts L‑DOPA to dopaquinone, which then polymerises spontaneously to eumelanin.
For heterologous expression in E. coli, the most efficient tyrosinase is MelA from Rhizobium etli (or from Streptomyces). This single enzyme can perform both hydroxylation and oxidation, simplifying the pathway to a one‑gene system. However, to ensure a sufficient supply of the precursor tyrosine, I co‑express tyrA (prephenate dehydrogenase), which diverts flux from the aromatic amino acid pathway towards tyrosine. The combination of tyrA and melA yields high melanin titres without the need for exogenous tyrosine supplementation.
Comparison of melanin types
Melanin type
Colour
Characteristics
Radioprotective relevance
Eumelanin
Black / brown
Indole polymer, high stability, broadband absorption
Used in this project
Pheomelanin
Red / yellow
Contains sulfur, less photostable
Moderate antioxidant
Pyomelanin
Reddish brown
Derived from homogentisic acid
Good metal chelator
Allomelanin
Variable
Nitrogen‑free, diverse structures
Potentially useful in extreme environments
2.2 The PprI Protein: Structure, Function, and Dual Utility
PprI (locus tag DR_0167 in D. radiodurans) is a 37 kDa protein consisting of three domains: an N‑terminal zinc‑peptidase domain, a helix‑turn‑helix (HTH) DNA‑binding domain, and a GAF‑like regulatory domain. Its primary biochemical activity is the cleavage of the repressor DdrO at a specific recognition site (ELRGKR). In living cells, this triggers the SOS‑like response, upregulating DNA repair enzymes.
For my inert composite, I am not using the protease activity; instead I exploit two side properties:
Extreme radiolytic stability – PprI retains its secondary structure (as measured by circular dichroism) after >10 kGy of gamma irradiation, whereas most proteins denature below 1 kGy.
ROS scavenging – The protein’s surface methionine and cysteine residues act as sinks for hydroxyl radicals and superoxide anions, protecting the surrounding matrix.
These properties make PprI uniquely suited as a protective additive for cellulose‑based composites. No other single natural protein combines such high radiation tolerance with antioxidant capacity.
The PprI protein is expressed with a six‑histidine tag (His‑tag) at the N‑terminus. This allows purification by affinity chromatography on a nickel‑NTA column, which retains His‑tagged proteins. After washing away impurities, the protein is eluted with imidazole. This method yields >90 % purity in a single step.
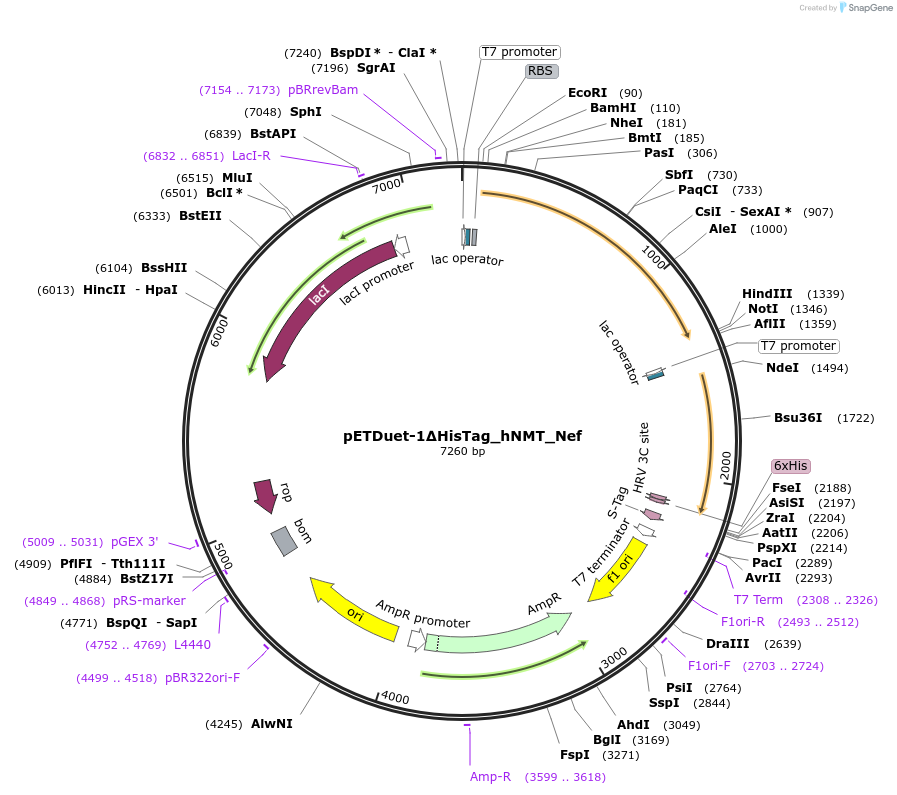
Figure 8: Map of the pETDuet‑1 vector. The two independent expression cassettes are shown with their T7 promoters, multiple cloning sites (MCS1 and MCS2) and T7 terminators. Cassette 1 contains the pprI gene; cassette 2 contains the tyrA‑melA operon.
2.3 Why E. coli BL21‑AI as the Production Host
The chosen production host is E. coli BL21‑AI (Thermo Fisher). This strain contains an arabinose‑inducible T7 polymerase gene inserted into the chromosome, providing extremely tight control of expression. Leaky expression of PprI (which can be toxic due to its protease activity) is minimised in the uninduced state. Upon addition of arabinose, T7 polymerase is produced and drives high‑level expression from the pETDuet‑1 T7 promoters. This inducible system also allows me to optimise the timing of protein and melanin production, separating growth phase from production phase.
3. AUTOMATION PLATFORM DESIGN (Aim 2)
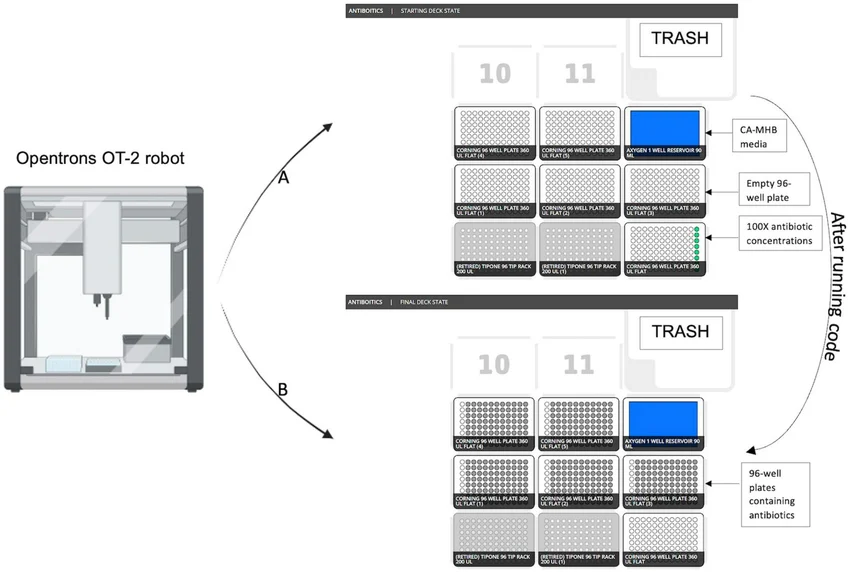
Although the project focuses on a single engineered strain, I still use automation to accelerate optimisation. The Opentrons OT‑2 robot is used to test dozens of culture conditions in parallel, eliminating manual bottlenecks.
3.1 Hardware Components
Opentrons OT‑2 robot with 96‑channel or 8‑channel pipette (depending on volume).
Temperature module for pre‑warming media and for holding plates during induction.
Magnetic module (optional) for cell separation after lysis.
Plate reader (e.g., BioTek Synergy HTX) for absorbance measurements (melanin at 405 nm, cell density at 600 nm).
Incubated shaker (set to 22 °C or 37 °C) for 96‑deep‑well plates.
Figure 9: Opentrons OT‑2 robot with robotic arm and 96‑well plate. This equipment automates pipetting, inoculation and sampling.
3.2 Software and Protocol Architecture
Python scripts control the robot. The workflow is modular:
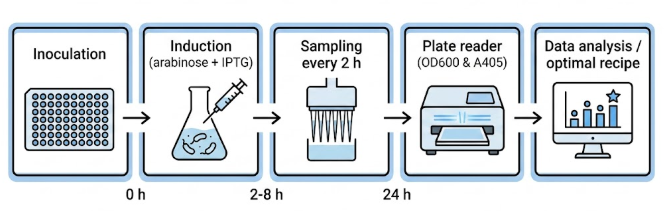
Inoculation module – automatically transfers overnight starter cultures into fresh 96‑well plates containing varying media compositions (LB, TB, or custom defined media with different carbon/nitrogen sources).
Induction module – adds IPTG (0.1–1.0 mM) and/or arabinose (0.02–0.2 %) at specified times (OD₆₀₀ ~0.6).
Copper supplementation – the tyrosinase MelA requires Cu²⁺ as a cofactor; the robot adds CuSO₄ (0.5–2.0 mM) to test its effect on melanin yield.
Sampling module – at regular intervals (2, 4, 6, 8 h post‑induction) transfers 200 µL of culture to a clear 96‑well plate for OD₆₀₀ (growth) and to a separate plate for melanin quantification (cells are pelleted by centrifugation; supernatant measured at 405 nm).
Harvest module – after 24 h, the robot adds NaOH (to solubilise melanin) and then HCl to precipitate it, collecting the precipitate by centrifugation. (Acid precipitation is done manually in a fume hood to protect the robot from corrosive fumes.)
The scripts use the official Opentrons API (opentrons.execute). Separate functions are defined for each module, and nested loops iterate over the different conditions of the experimental design. Communication with the plate reader is done via HTTP commands or shared CSV files. All code is version‑controlled on GitHub to ensure reproducibility.
Figure 10: Automated workflow: (1) culture preparation, (2) induction with arabinose/IPTG, (3) periodic sampling, (4) absorbance reading on plate reader, (5) data processing.
3.3 Experimental Workflow for Optimisation
A Design of Experiments (DoE) approach is used to vary three factors (IPTG concentration, temperature, Cu²⁺ concentration) at three levels each (27 conditions). Each condition is tested in triplicate on a single 96‑well plate, with positive control (known melanin producer, e.g., Streptomyces) and negative control (empty vector). The robot runs the entire plate in 24 h, collecting growth and melanin data every 2 h. The optimal condition is defined as the one giving the highest melanin absorbance normalised by final OD₆₀₀ (specific yield).
Figure 11: Example of a 96‑well plate after induction. Darker wells indicate higher melanin production. The colour gradient reflects the different conditions tested.
4. ASSAY DEVELOPMENT
4.1 Melanin Quantification
Melanin absorbs strongly at 405 nm with a featureless spectrum. To quantify concentration, I first pellet cells, measure supernatant A₄₀₅, and subtract a blank (media only). A standard curve is prepared using synthetic eumelanin (Sigma‑Aldrich) dissolved in 0.1 M NaOH. For absolute quantification, I dry a known volume of purified melanin and weigh the residue (gravimetric analysis), then use that to calibrate the A₄₀₅ readings.
4.2 PprI Characterisation
After culturing under optimal conditions, I lyse cells by sonication or chemical lysis (BugBuster), and purify PprI via its N‑terminal His‑tag using Ni‑NTA spin columns (manual step after robot harvest). The eluate is then analysed by:
SDS‑PAGE – a single band at ~37 kDa indicates purity >90 %.
LC‑MS (intact mass) – on a Waters Xevo G3 QToF, the observed mass must match the theoretical mass of the His‑tagged PprI (calculated from the codon‑optimised sequence). A deviation <50 ppm confirms correct expression.
LC‑MS/MS peptide mapping – tryptic digest followed by tandem MS confirms the amino acid sequence covering at least 80 % of the protein.
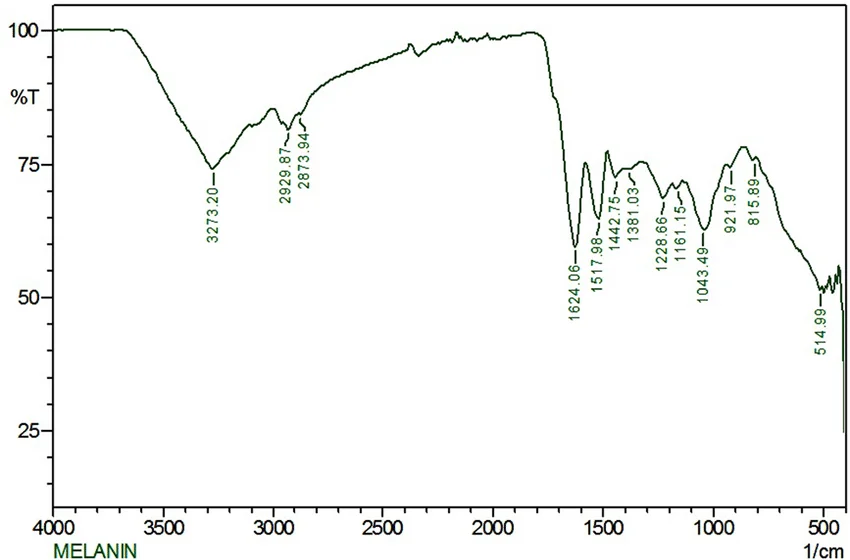
Figure 12: FTIR spectrum of purified melanin. Characteristic peaks are at 3400 cm⁻¹ (OH/NH), 1620 cm⁻¹ (aromatic C=C) and 1200 cm⁻¹ (C‑OH).
Figure 13: Coomassie‑stained SDS‑PAGE gel. The lane shows a single band at 37 kDa, indicating high purity of PprI.
4.3 Composite Fabrication and Gamma Testing (Aim 3)
Cellulose sheets (Whatman No. 1 filter paper, 0.2 mm thick) are cut into 5 × 5 cm squares. Purified melanin (10 mg/mL in 0.1 M NaOH) and PprI (2 mg/mL in PBS) are mixed at a 5:1 (w/w) ratio to give a total coating concentration of 20 mg per square centimetre. The mixture is spray‑coated evenly onto both sides of the cellulose and dried overnight at room temperature.
The 5:1 ratio was chosen based on preliminary studies indicating that too high a protein concentration interferes with melanin adhesion, while too low a concentration does not provide sufficient radical capture. This ratio will be further optimised in future experiments using a new DoE.
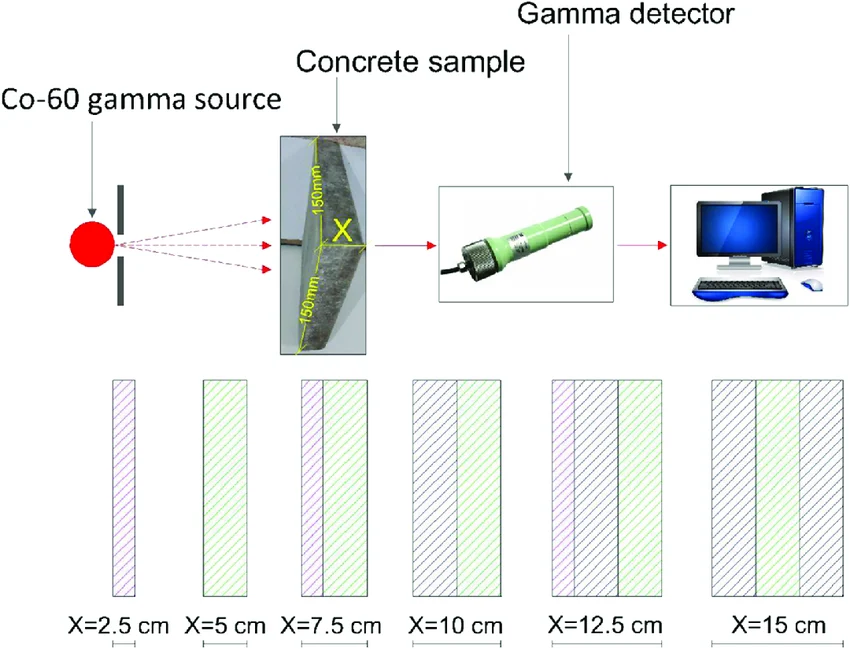
The coated and uncoated (control) samples are individually placed in front of a calibrated dosimeter (thermoluminescent or semiconductor) and exposed to a Cobalt‑60 gamma source (1.25 MeV average energy) at a dose rate of ~1 Gy/min. The total dose is 10 Gy (simulating one week of deep‑space exposure). The dosimeter reading behind each sample is recorded, and the linear attenuation coefficient μ is calculated using the Lambert‑Beer variant:
I = I₀ · e^{-μx}
where I is the transmitted dose, I₀ the incident dose, and x the thickness (0.2 mm cellulose plus coating). For the uncoated control, the coating contribution is treated as zero.
The attenuation coefficient of lead at the same energy is ~0.22 cm⁻¹; a BioShield coefficient of 0.15 cm⁻¹ (70 % of lead) at one‑tenth the weight would be considered a major success.
Figure 14: Experimental setup for gamma attenuation measurement. The Cobalt‑60 source emits radiation that passes through the coated cellulose sample; a dosimeter records the transmitted dose.
5. EXPECTED OUTCOMES AND APPLICATIONS
5.1 Deliverables
Genetic construct – a complete Benchling‑designed plasmid map (pETDuet‑1::tyrA‑melA‑pprI) with annotated features, plus codon‑optimised sequence files (FASTA, GenBank).
Automated protocol – an open‑source Python script for the Opentrons OT‑2 that implements the DoE screening, including data logging and analysis.
Optimised production conditions – a table of the best temperature, IPTG, and Cu²⁺ concentrations for maximum melanin yield (mg/L/OD).
Characterised biomolecules – FTIR spectra confirming eumelanin structure, LC‑MS data confirming PprI mass and sequence, and SDS‑PAGE gel images showing purity.
Radiation test results – attenuation coefficients for coated vs. uncoated cellulose, plotted with error bars, and a comparison to lead at the same areal density.
Expected results table
Parameter
Expected value
Measurement method
Melanin concentration (mg/L)
> 500 mg/L
Absorbance at 405 nm (standard curve)
PprI purity
> 90 %
SDS‑PAGE (densitometry)
PprI mass
37 kDa ± 50 ppm
Intact LC‑MS
PprI sequence coverage
> 80 %
LC‑MS/MS (peptide mapping)
Attenuation coefficient (μ)
0.15 cm⁻¹
Lambert‑Beer law with Co‑60 source
Coating weight
20 mg/cm²
Gravimetry
5.2 Applications
Aerospace – lightweight, flexible shielding for spacecraft, habitats, and extravehicular activity suits. The ability to produce the coating in‑situ using local biomass (e.g., cellulose from plant waste) aligns with NASA’s in‑situ resource utilisation (ISRU) roadmap.
Nuclear facilities – biodegradable paints and sealants that provide supplemental protection to workers and can be disposed of without heavy metal contamination.
Medicine – topical melanin‑PprI creams could protect skin during radiation therapy; the PprI component may reduce oxidative damage.
Consumer products – natural melanin‑based sunscreens that are biodegradable and reef‑safe.
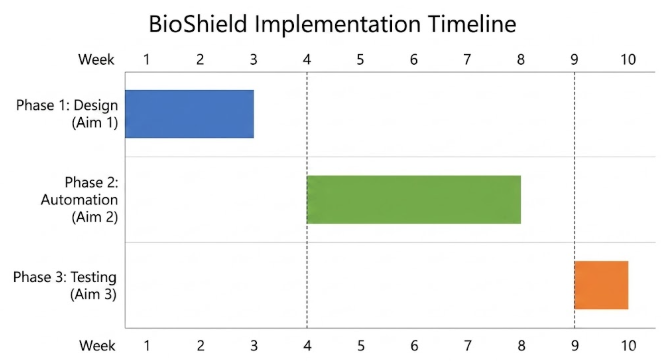
6. IMPLEMENTATION PLAN
6.1 Phase One: Digital Design (Aim 1) – Weeks 1‑3
Week 1 – Obtain sequences: pprI (DR_0167 from D. radiodurans), tyrA (DR_1935), melA (from R. etli CP000137). Import into Benchling.
Week 2 – Codon‑optimise all three genes for E. coli BL21‑AI using Benchling’s tool; check for internal restriction sites that interfere with cloning.
Week 3 – Simulate Gibson assembly of three fragments (pprI into MCS1; tyrA‑melA operon into MCS2 of pETDuet‑1). Generate final plasmid map, order the synthetic construct from Twist Bioscience.
Week 4 – Receive construct, transform into E. coli BL21‑AI, verify by colony PCR and restriction digest.
Week 5 – Write and debug Opentrons Python scripts for inoculum preparation, 96‑well plate setup, and automated sampling.
Week 6 – Run DoE screening (27 conditions, triplicate) with the robot; collect growth and melanin data.
Week 7 – Analyse data to identify optimal conditions; perform a confirmation run in shake flasks.
Week 8 – Scale‑up to 1 L cultures under optimal conditions; harvest cells by centrifugation, lyse, and purify PprI via Ni‑NTA; extract melanin by acid precipitation.
Week 9 – Characterise purified melanin by FTIR (compare to reference eumelanin spectrum). Characterise PprI by SDS‑PAGE, LC‑MS intact mass, and LC‑MS/MS peptide mapping.
Week 10 – Coat cellulose sheets, expose to Cobalt‑60 gamma source, measure attenuation with dosimeter; calculate linear attenuation coefficient; compare to lead reference.
Figure 15: Ten‑week project timeline showing the main phases and milestones.
7. RESOURCE REQUIREMENTS
7.1 Equipment
Opentrons OT‑2 robot (used) – $5,000
Plate reader (UV‑Vis) – lab shared resource
Shaking incubator (30 °C and 37 °C) – lab shared
Centrifuge (for 50 mL tubes and 96‑well plates) – lab shared
FTIR spectrometer – access via chemistry department
LC‑MS (Waters Xevo G3 or BioAccord) – access via Waters academic program
Cobalt‑60 gamma source – external facility (e.g., university irradiation center)
Note: The budget reflects a transition to TRL 3‑4. Costs for additional Opentrons modules (Heater‑Shaker, Temperature Module) are excluded but recommended for Aim 2.
Figure 16: Logos of key industry partners and suppliers for the project (Twist Bioscience, Opentrons, Thermo Fisher, Millipore Sigma, Waters).
8. RISK ASSESSMENT AND MITIGATION
8.1 Technical Risks
Low yield of melanin – the tyrosinase may require specific copper loading or oxygen availability. Mitigation: test Cu²⁺ concentration from 0.1‑5 mM; increase aeration by using high‑speed shaking (300 rpm) and deep‑well plates with gas‑permeable seals.
PprI toxicity – leaky expression may kill cells before induction. Mitigation: use BL21‑AI strain (arabinose inducible) and maintain cultures with 0.2 % glucose to repress T7 polymerase.
Poor solubility of PprI – inclusion bodies may form. Mitigation: reduce induction temperature to 16 °C; add 0.5 M sorbitol and 2.5 mM betaine to the medium; use a milder lysis buffer with 0.5 % Triton X‑100.
Cellulose degradation during coating – alkaline melanin solution may weaken the paper. Mitigation: neutralise the melanin solution with Tris‑HCl (pH 7.0) before coating; apply multiple thin layers instead of one thick layer.
Cross‑contamination in the robot – melanin can stick to pipette tips and cause well‑to‑well contamination. Mitigation: use fresh tips for each transfer and program acid washes (0.1 M HCl) between samples.
8.2 Biological Risks
The engineered E. coli expresses a protease (PprI) that could theoretically interfere with host cell metabolism, but the strain is non‑pathogenic and contained at BSL‑1. All waste is autoclaved before disposal. No antibiotic resistance genes beyond kan‑R are used, and the construct is not mobilised. Extraterrestrial contamination is not an issue at this terrestrial proof‑of‑concept stage.
8.3 Timeline Risks
Gene synthesis from Twist usually takes 2‑3 weeks; ordering early mitigates delays. Opentrons programming may require debugging; I allocate an extra week for script validation. Access to the Cobalt‑60 source may be limited; I have identified a backup facility at a local cancer hospital (linear accelerator) that can simulate gamma exposure.
9. CONCLUSIONS
BioShield demonstrates that a lightweight, biologically‑produced radioprotective composite can be engineered using accessible synthetic biology and automation tools – specifically, the melanin pathway enzymes (TyrA and MelA) together with the PprI bio‑stabiliser, expressed from a single pETDuet‑1 construct in E. coli, optimised via Opentrons‑driven DoE, and validated by gamma‑attenuation measurements. The expected linear attenuation coefficient (0.15 cm⁻¹) provides 70 % of lead’s performance at a fraction of the mass, potentially reducing launch costs for deep‑space missions. Moreover, the open‑source design and automated protocol allow for rapid adaptation to other melanin‑producing strains or other protective biomolecules, making BioShield a versatile platform for biological radiation protection. Future work will focus on scaling the biomanufacturing process and testing the composite under simulated space conditions (thermal vacuum, heavy‑ion radiation, atomic oxygen erosion). The ultimate goal remains in‑situ resource utilisation: using microorganisms growing on local biomass to produce radiation shields on the Moon or Mars, thereby liberating human exploration from the tyranny of the rocket equation.
Figure 17: Conceptual view of a Mars habitat with melanin‑coated domes, produced in situ via biomanufacturing (ISRU).
Appendix: 3‑Minute Presentation Overview
The following three slides summarize the BioShield project as presented in the final 3-minute defense. Each slide includes a brief verbal script and the key visuals of what I had to mention that day. Obviously, there were some things I couldn’t say because of nerves AJAJA.
Slide 1 – The Problem: Why We Need a Better Shield
Verbal script “Radiation is a serious problem. In space, cosmic rays damage DNA and cause cancer. A trip to Mars would give astronauts a dangerous dose. On Earth, nuclear workers and radiotherapy patients suffer similar tissue damage. The usual solution is lead – but lead is extremely heavy. Launching one kilogram into space costs tens of thousands of dollars. Worse, when cosmic rays hit lead, they create dangerous secondary radiation. So how do we protect people without carrying heavy metals? I turned to biology.”
Slide 2 – Aim 1: Engineering the Biological Shield
Verbal script “My first aim is to design a genetic blueprint. I use two natural molecules: melanin, which absorbs radiation and turns it into heat, and the protein PprI from Deinococcus radiodurans, which captures free radicals. I assemble three genes – tyrA, melA (melanin) and pprI – into a dual‑promoter plasmid called pETDuet‑1. The two independent T7 promoters act as separate switches, so the melanin factory and the protein factory run in parallel without interfering. I simulate Gibson Assembly and optimise the DNA letters for E. coli. The final digital file is ready for synthesis.”
Slide 3 – Aim 2, Aim 3 and Conclusion
Verbal script “Aim 2 automates production. An Opentrons robot tests 96 combinations of temperature, inducer, and copper to find the recipe that maximises melanin and PprI. After purification, I confirm melanin by FTIR and PprI by LC‑MS and SDS‑PAGE. Aim 3 tests the shield: I coat ordinary cellulose paper with the purified biomolecules and expose it to a Cobalt‑60 gamma source. A dosimeter measures how much radiation passes through. The expected result is that this thin biological coating blocks 70 % of the gamma rays – almost as well as lead, but at a fraction of the weight. BioShield proves that biology can outperform heavy metals. It is lighter, safer, and could even be grown on Mars from local resources. Thank you.”
Group Final Project
Computational Engineering of the MS2 Lysis Protein to Improve Stability, Titers, and Toxicity
After reviewing the provided literature on the MS2 lysis protein (L) and discussing the project aims, our group has decided to focus on three interconnected goals:
Goal 1: Increase the stability of the L protein
As the “easiest” goal, it is the most computationally tractable. A stabilized protein is less prone to degradation and misfolding, which could directly lead to higher functional titers and serve as a robust starting point for any subsequent engineering.
Goal 2: Increase bacteriophage titers through improved lysis efficiency.
Phage therapy relies on high phage titers for effective bacterial killing and scalable manufacturing, but phage production can be limited by inefficient lysis or poor coordination between phage replication and host destruction. Improving the efficiency and timing of host cell lysis can therefore directly increase the number of phage particles released per infected cell.
The MS2 L protein is a small 75–amino acid membrane protein that triggers bacterial lysis and is essential for the release of new phage particles. In the paper Mutational analysis of the MS2 lysis protein L, it is described how MS2 L functions as a single-gene lysis protein that disrupts bacterial cell envelope integrity without classical enzymatic activity. Additionally, L interacts with the host chaperone DnaJ, which modulates its activity and timing of lysis. In MS2 Lysis of Escherichia coli Depends on Host Chaperone DnaJ it is shown that lysis timing strongly affects the number of virions produced before the host cell bursts, meaning that engineering improved L variants may increase overall phage titers.
Goal 3: Increase the toxicity of the lysis protein.
This proposal addresses the subproblem of increasing the toxicity of the L lysis protein from Bacteriophage MS2. Instead of random mutagenesis, toxicity will be approached as a multi-factor optimization problem involving structural stability, membrane insertion, oligomerization efficiency, and expression kinetics in Escherichia coli. The objective is to design L variants that enhance membrane disruption while maintaining proper folding and stability.
E. coli chaperone DnaJ.
Additionally, we will explore disrupting the interaction between the L protein and the E. coli chaperone DnaJ.
The reading “Identification MS2 lysis protein dependency on DnaJ” establishes this interaction as critical for function. By computationally predicting and then disrupting this interface, we can test its necessity and potentially create a DnaJ-independent lysis mechanism, offering a new avenue for controlling lysis timing.
Together, these three goals form a coherent strategy: stabilizing the L protein may improve its folding and expression, which can increase functional titers, while further engineering of membrane disruption and host interactions may increase toxicity and lysis efficiency.
Proposed Computational Tools and Approaches
Proposed Tools and Approaches We will build a computational pipeline using the tools introduced in recitation and the provided resources. The key steps and tools are:
Step 1: Structural Modeling of the L Protein
Tool: AlphaFold2 (via ColabFold for ease of use).
Why: No high-resolution experimental structure of the full-length MS2 L protein exists. A reliable 3D model is the absolute foundation for all downstream analysis, allowing us to visualize which parts are structured vs. disordered.
Step 2: Modeling the L-DnaJ Complex
Tool: AlphaFold-Multimer.
Why: To disrupt the interaction, we first need to know where it occurs. AlphaFold-Multimer is the current state-of-the-art for predicting protein-protein complexes and will generate a testable model of the L protein bound to E. coli DnaJ.
Step 3: In Silico Mutagenesis for Stability
Tool: Rosetta (or FoldX). Specifically, the ddg_monomer application for predicting changes in folding free energy (ΔΔG).
Why: These tools are parameterized using vast amounts of experimental data on protein stability. They can systematically mutate each residue in our L protein model and predict whether the change (e.g., A->V) makes the protein more stable (negative ΔΔG) or less stable (positive ΔΔG).
Step 4: Visualizing and Selecting Interface Mutations
Tool: PyMOL and the HTGAA Protein Engineering Tools spreadsheet.
Why: We will use PyMOL to visually inspect the predicted L-DnaJ complex from Step 2 and select residues at the interface. We will then use the spreadsheet to check the conservation of those residues and manually design mutations (e.g., swapping a large hydrophobic residue for a charged one) predicted to break the interaction.
Protein Language Models (PLMs)
Protein language models such as ESM or ProtBERT will be used to perform in silico mutagenesis on the MS2 L protein sequence. These models can suggest mutations that preserve structural and functional constraints learned from large protein datasets.
This approach allows us to generate multiple candidate mutations across the L protein, avoid mutations likely to disrupt folding, and explore sequence space beyond naturally occurring variants
AlphaFold Structure Prediction
Each candidate L variant will be analyzed using AlphaFold to predict protein structure and membrane topology. Since the C-terminal transmembrane region is essential for lytic activity, structural prediction will help identify mutations that preserve this functional domain.
Structural predictions will also help identify:
misfolded variants
mutations that destabilize the transmembrane region
variants that may alter oligomerization or membrane insertion
Interaction Modeling with Host Proteins
Because MS2 L interacts with the DnaJ chaperone, which affects lysis timing, candidate variants can be evaluated using AlphaFold-Multimer to predict changes in the L–DnaJ interaction.
This could help identify variants that:
maintain necessary folding assistance
reduce excessive dependency on host chaperones
improve robustness of lysis across physiological conditions
Proposed Computational Strategy
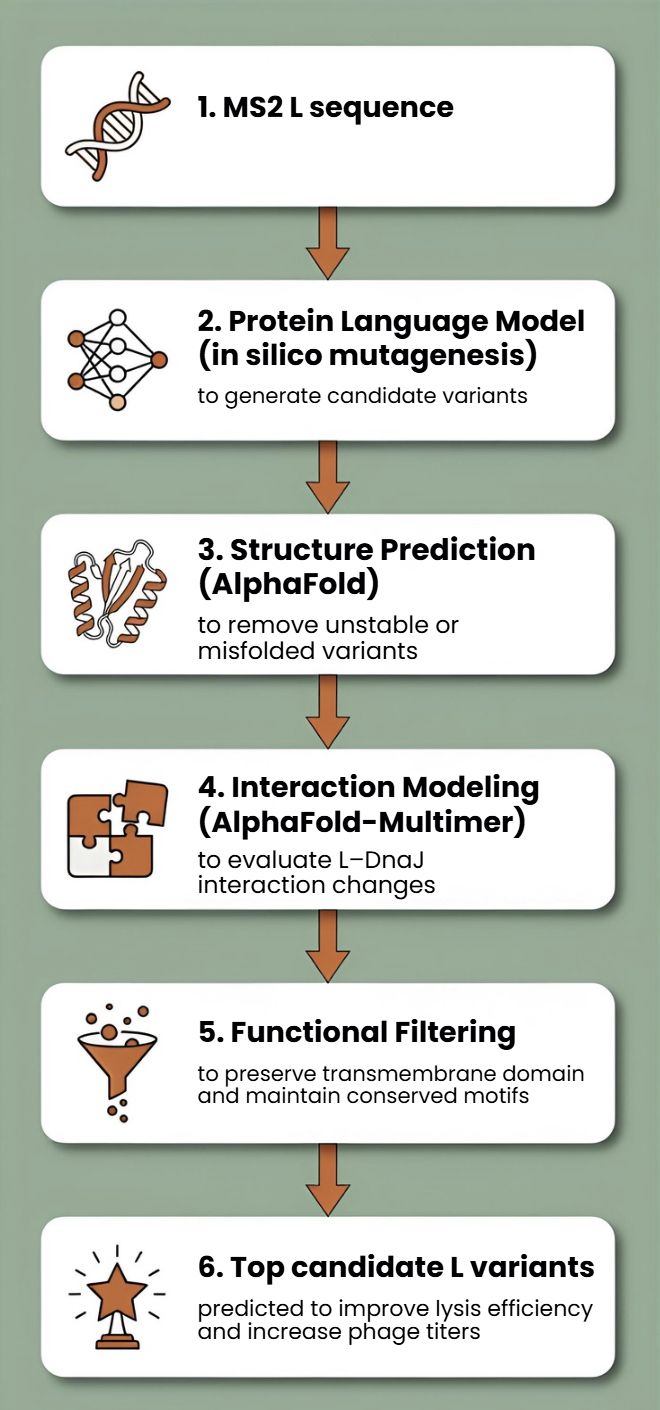
First, protein language models (e.g., ESM-2, ProtT5) will be used to perform directed in silico mutagenesis. These models capture evolutionary constraints and residue interactions, enabling the generation of structurally plausible variants while identifying mutation-tolerant and functionally critical positions. This step efficiently reduces the combinatorial search space.
Second, predicted variants will be structurally evaluated using AlphaFold2 for monomer folding and AlphaFold - Multimer to assess oligomerization and interaction with host factors such as DnaJ.
Third, membrane compatibility will be analyzed using membrane-aware modeling (RosettaMP) and selected molecular dynamics simulations.
Fourth, ΔΔG prediction tools (e.g., FoldX, Rosetta energy functions) will filter out destabilizing mutations.
In parallel, codon optimization algorithms will redesign selected variants for improved expression in E. coli, as toxicity depends on both structure and intracellular concentration.
Potential Pitfalls
Pitfall 1: Dynamic Regions and Model Quality
The L protein is small and likely has flexible/disordered regions, especially in its N-terminal domain.
Pitfall 2: Stability vs. Function Trade-off
A mutation that makes the protein more stable in its monomeric state might prevent it from undergoing the necessary conformational changes to oligomerize and form a pore in the membrane.
Pitfall 3: Lack of Membrane Context
Our stability predictions (Rosetta) are performed in a virtual “aqueous” environment and do not account for the energetic complexity of the lipid bilayer.
Limited biological data: There is still limited structural and mechanistic knowledge about MS2 L.
Cellular context not captured computationally Protein modeling tools may not fully capture membrane environment.
One limitation is the scarcity of quantitative datasets linking specific mutations to measured lysis kinetics.
L-Protein Mutants
To generate the first two mutations in the L protein of bacteriophage MS2 within the transmembrane region, I selected the top candidates predicted by the Python models and the spreadsheet analysis for that region. I applied the same approach to the soluble region, ensuring that all mutations were introduced at amino acid positions with less constrained mutability.
METRFPQQSQQTPASTNRRRPFKHEDYPCRRNQRSSTLlVLIFLAIFLSlFTlQLLLSLLEAVIRTVTTLQQLLT
METRFPQQSQQTPASTNRRRPFKHEDYPCRRNQRSSTLheLnlvpnFLleFTNQLhLSLLEAeIRTVTTLQQLLT
METRqPQQqQQTPASTNRRRPFKHEDYPrRRNQRSSTLYVLIFLAIFLSKFTNQLLLSLLEAVIRTVTTLQQLLT
lEiRqPQQqQQTPASTNRRRPFKHEDYPrRRNQRSSTLYVLIFLAIFLSKFTNQLLLSLLEAVIRTVTTLQQLLT
For the final mutation, which was the most aggressive, I introduced mutations in both regions across all possible amino acid positions.