Long version: Software Developer with hands-on experience in systems programming, test automation, and
distributed infrastructure. Skilled in C, C++, Python, and full-stack development across networking, cloud platforms,
and databases. Architected and maintaining OLIVIA: a self-hosted distributed system demonstrating networking
concepts at scale. Experienced in QA and debugging production systems. Works effectively with remote international
teams. Passionate about leveraging AI in dev workflows, infrastructure automation, and networking solutions.
Assignment 1 Background Malaria is a dangerous disease that affects hundred of millions of people worldwide. Each year, there are almost 300 million cases, with 600,000 deaths, 95% of which are disproportionately in Africa.
The only known cure for malaria for the longest time was quinine, a drug that was used to treat the disease in the 19th century. In the 1970s, Tu Youyou discovered artemisinin, a compound that is more effective in eradicating malaria than quinine. A few years later, she was able to synthesize it on a much larger scale using a modified yeast cell.
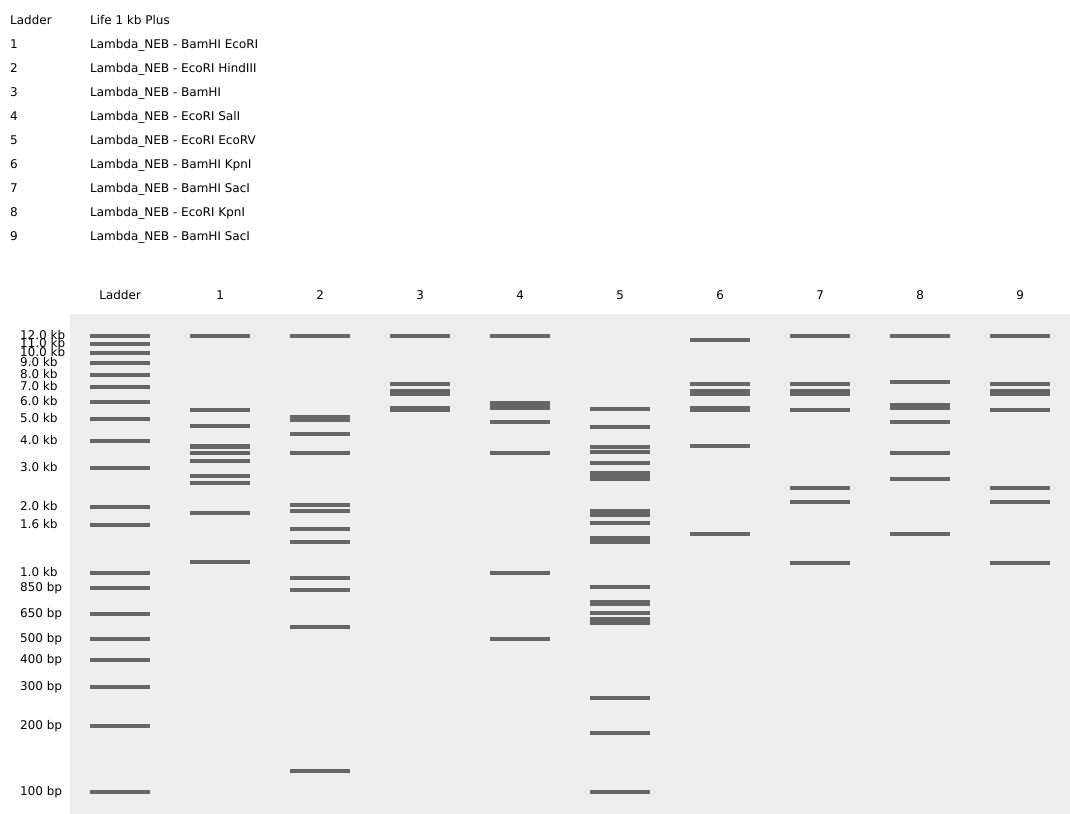
Part 1: Benchling & In-silico Gel Art placeholder text
Part 3: DNA Design Challenge 3.1: Choose your protein I decided to pick the Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1). This is a large family of functionally different proteins that are created from a group of about 60 genes that the Plasmodium falciparum parasite can turn on and off to evade the immune system. I decided to choose one of these proteins for analysis as I want to understand the mechanism of this one protein individually to begin to understand the many different ways a malarial infection can occur within the human body.
Subsections of Homework
Week 1 HW: Principles and Practices
Assignment 1
Background
Malaria is a dangerous disease that affects hundred of millions of people worldwide. Each year, there are almost 300 million cases, with 600,000 deaths, 95% of which are disproportionately in Africa.
The only known cure for malaria for the longest time was quinine, a drug that was used to treat the disease in the 19th century. In the 1970s, Tu Youyou discovered artemisinin, a compound that is more effective in eradicating malaria than quinine. A few years later, she was able to synthesize it on a much larger scale using a modified yeast cell.
I personally caught malaria in 2007 and my father caught dengue in Vietnam in 2005. They are very common diseases that are largely eradicated nowadays thanks to health policies and an improvement in quality of life.
Malaria, however, is adaptive. In 2008, reports came out from Southeast Asian countries that the parasite was evolving to be resistant to artemisinin. More reports came out in 2009, 2011, and 2012.
There are also reports in Vietnam in 2024 that the Rapid Detection Test (RDT) may no longer be effective as the parasite evolved to not express the 2 main proteins that the RDT is built to detect. Meaning, malaria is slowly becoming underdiagnosed and proper diagnoses will require more expensive methods that are not available in many countries.
The rise in resistance is a pain point for the world as many countries’ eradication efforts are now hampered by what is simply selective pressure. This threat must be addressed before the new strains reach the most vulnerable populations, specfically Africa.
Project Goals
There are 5 main species of parasites from the Plasmodium genus that causes malaria in humans, most prevalent being P. falciparum on the African continent and P. vivax outside of Sub-Saharan Africa. We will be focusing on P. falciparum.
For my project, I want to use biological circuitry to create a cure that is also adaptive to the disease. By analyzing the evolutionary mechanism of the parasite, we can synthesize products that can target our treatments to work around its defensive mechanisms.
Also, to adapt to the ever-changing nature of the parasite, I would like to create a circuitry that can adapt to whatever ingredients we feed to cheaply and quickly synthesize new, targeted drugs.
Having a simple-to-replicate medication that can be modified to target different strains of malaria or to target different weaknesses of P. Falciparum will drastically improve life expectancies and help curb malaria deaths worldwide.
Policy Goals
For this biological circuitry idea to work, we will need some ground rules. The design, use, and proliferation of a modified cell containing synthetic circuitry could prove dangerous in many ways. It could either affect the creator, distributor, consumer, or the environment.
Thus, strict emphasis must be placed on safety and containment.
On top of that, the whole point is to make malarial treatments more readily available. Thus accessibility in all its definitions should be considered. Below are some I have come up with:
Biosafety
Make sure the modified cells will not have pathogenic/virulent factors that might cause new disease
Ensure containment of bacteria in safe environment
Ensure generated product and byproducts are not harmful to patients
Accessibility
Ensure affordability
Preservability & Procurement
Actions and Actors
Researchers
Action 1 – Ensure modified cells are not pathogenic/virulent
Creating modified cells comes with a risk of misuse or miscreating a dangerous pathogen. It is important the modified cell does not post any risk at all to the creator, distributor, and user of its products.
Purpose: To not design a biohazard for the community
Design: Test of pathogenicity before starting the bioreactor
Assumptions: The test are enough to ensure no pathogenicity, this is maintained stable in the bioreactor
Risks of Failure & “Success”:
Failure: there is pathogenicity, people can be sick, directly or indirectly.
Success: No pathogens.
Action 2 – Ensure generated product and byproducts are not harmful to the patient
Purpose: Elaborate innocuous products that can be used as medicine
Design: Previous test in different biological models such as murine models and appropriate clinical trials
Assumptions: The effects of the byproducts will not variate between people
Risks of Failure & “Success”:
Failure: Unexpected outcomes in different patients, potentially dangerous.
Industrial plant designer
Action 3 – Ensure containment of modifed cells in safe environment
Purpose: Prevent cell leakage and contact with the environment
Design: Appropriate infrastructure for biorreactors and checkpoints to ensure appropriate fluid levels
Assumptions: Good practices from the operators in the industrial plant
Risks of Failure & “Success”:
Failure: leakage that affects the surrounding environment.
Success: Very elaborate plant design that is not affordable
Local government and regulating bodies
Action 4 – Government subsidies and support
Derivatives of artemisinins are generally stable in room temperature, but will quickly deteriorate under high humidity and the extreme heat of Africa, South and Southeast Asia. Furthermore, the cost of each dose could range from cents to hundreds of dollars. Government subsidies can help keep prices low (like in Southeast Asia) and proper storage and transport can protect medication stock.
Purpose: Make medicine affordable and widely available to vulnerable communities
Design: Price controls through public subsidies and partnerships
Assumptions: The government will have the budget to invest in health
Risks of Failure & “Success”:
Failure: Medicine still not affordable to low income families.
Success: High subsidies might lead to fake products.
Scoring
After realizing too late that I’ve used the scale backwards, the scoreboard actually has 3 as the most important and 1 as the least important.
In other words, higher scores => more importance for this specific score board.
Tip
This is a funny case of Sienna not being able to read properly. Keep count until the end of term because this WILL happen again. My over-under is 7.5+ times.
Actors
Researchers
Industrial plant designers
Local governments
Does the option
Action 1
Action 2
Action 3
Action 4
Ensure Biosafety
• Ensure modified cells are not pathogenic/virulent that might cause new diseases
3
3
n/a
n/a
• Ensure containment of bacteria in safe environment
1
2
3
n/a
• Ensure generated product and byproducts are not harmful to patients
3
3
1
n/a
Accessibility
• Ensure affordability
n/a
n/a
n/a
3
• Preservation & procurement
n/a
n/a
n/a
3
Priorities
Using the score board, it can be seen that in order of importance, Action 2 takes the highest precedent. Action 1 can be argued to be more important as a deadly disease spreading everywhere is an issue anywhere. However, due to some oversight, I just now realized that Action 2 technically encompasses parts of Action 1, in the sense that a patient should not be harmed.
I will argue that is the primary goal of the project. The idea of synthesizing an adaptable product is to cure people, thus, harming people is antithetical to that and utmost priority must be placed on such a duty. Subsidies and storage are important to manipulating and keeping drug prices low, but I don’t think it can take priority over the main utility of the project: curing disease.
Once that is done, we can begin worrying about logistics.
Week 2 Lecture Prep
For Professor Jacobson
Question 1
DNA polymerases are highly accurate but not perfect. Their intrinsic error rate is roughly one mistake per million nucleotides added (10-6). When this number is compared with the size of the human genome-about 3.2 billion base pairs-it becomes clear that thousands of errors would appear if no additional safeguards existed.
To visualize this, copying the human genome with that error rate would be similar to transcribing a multi-billion-letter manuscript and ending up with a few thousand typos spread across the entire text.
Living systems compensate for this mismatch using several layers of quality control. During DNA synthesis itself, polymerases can remove incorrect bases via a 3' -> 5' exonuclease proofreading activity. Errors that escape this first checkpoint are later corrected by post-replication mismatch repair pathways, such as the MutS system, which scans the DNA and fixes mispaired bases.
Question 2
Coding capacity of DNA for a human protein
Because the genetic code is redundant, a single protein sequence can, in theory, be encoded by an enormous number of distinct DNA sequences. A typical human protein is around 345 amino acids long (≈1,036 bp). Since most amino acids are specified by multiple synonymous codons, the number of possible DNA sequences that could encode the same protein quickly becomes astronomical.
If each amino acid had, on average, three possible codons, the total number of theoretical coding sequences would be on the order of 3³⁴⁵ (approximately 10165).
However, only a small subset of these sequences is biologically or technically viable. Factors that limit usable coding options include:
Extreme GC or AT content, which promotes problematic secondary structures.
Sequence motifs that destabilize mRNA or promote cleavage.
Codon usage preferences, which vary between organisms and affect translation efficiency.
Long repeats or homopolymers, which complicate synthesis and assembly.
Local thermodynamic constraints, such as unfavorable base pairing energies.
For Dr. LeProust
Question 1
The dominant method for making synthetic oligonucleotides today is phosphoramidite chemistry, introduced in the early 1980s by Caruthers. Despite its age, it remains the foundation of nearly all commercial DNA synthesis platforms.
Question 2
As oligos become longer than ~200 nucleotides, the fraction of incomplete or truncated products increases significantly. Although companies like Twist have demonstrated unusually long directly synthesized fragments (up to ~700 nt), these remain exceptions rather than the norm.
Question 3
Synthesizing a 2 kb gene directly is impractical. Instead, long genes are produced by assembling many shorter oligonucleotides, reducing cumulative error rates and increasing overall accuracy.
For Dr. George Church
Question 1
Animals require ten essential amino acids from their diet: histidine, isoleucine, leucine, lysine, methionine, phenylalanine, threonine, tryptophan, valine, and arginine.
This fact directly undermines the idea of a “lysine contingency.” Since animals are already incapable of synthesizing lysine, removing lysine biosynthesis does not meaningfully restrict survival. Carnivores obtain lysine from prey, while herbivores consume lysine-containing plants, making lysine universally available within natural food webs. Effective biocontainment strategies must therefore rely on dependencies that do not exist in nature, such as non-standard amino acids or extensively recoded genetic systems, as discussed in Church’s work.
Question 2
Several biological coding systems already exist: DNA–DNA pairing through Watson–Crick rules, DNA–amino acid relationships via the genetic code, and amino acid–DNA recognition in systems like TALEs.
For amino acid–amino acid interactions, a plausible and intuitive “code” could be built from fundamental biochemical properties rather than strict sequences:
Electrostatic complementarity, where basic residues (K, R, H) interact with acidic ones (D, E).
Hydrophobic matching, in which nonpolar residues (L, I, V, F, W, A) preferentially associate.
Polar and hydrogen-bond interactions, involving residues such as S, T, N, Q, and Y.
Together, these principles could form a simple but robust framework for AA:AA recognition.
Question 3.
Sketch Response to Smart Red Blood Cells (Smart-RBC) Program
I propose engineering RBCs with genes from freeze-tolerant organisms like wood frogs (Rana sylvatica) and hibernating mammals that survive extreme cold and low oxygen without cellular damage. Wood frogs naturally produce antifreeze proteins and cryoprotectants (like glucose) that prevent ice crystal formation during winter freezing, while hibernating animals express protective chaperones that maintain function during hypoxia. By inserting these genes into RBC precursor cells before enucleation, the resulting Smart-RBCs would retain protective proteins while eliminating genetic transfer risks. This would temporarily enhance warfighters’ cold tolerance and oxygen delivery in Arctic operations, high-altitude missions, or extreme survival scenarios—without the weeks needed for natural acclimation. Since mature RBCs are enucleated and only last ~120 days, the protection is temporary and safe, providing exactly the performance boost needed when it matters most.
I decided to pick the Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1). This is a large family of functionally different proteins that are created from a group of about 60 genes that the Plasmodium falciparum parasite can turn on and off to evade the immune system. I decided to choose one of these proteins for analysis as I want to understand the mechanism of this one protein individually to begin to understand the many different ways a malarial infection can occur within the human body.
The sequence of the protein I chose is as shown below:
Maybe I should do a dropdown or callout to hide this ginormous block, huh? Or maybe you guys can have some fun scrolling like I did 👍
3.3 Codon Optimization
For this section, I will use Satisfactory and certain coding concepts to explain codon optimization as that was how I understood it.
Codon Optimization can be done due to genetic code degeneracy, where the same amino acid can map to different codons. In a situation where we implement one species’ genetic code onto another species, say manufacturing insulin using E. coli, the amino acids that is used in the original species may choose to bind with a specific codon which the E. coli may not have in abundance. By optimizing the codons, we “change the recipe” to make it bind with more abundant codons and thus speed up the process.
In Satisfactory, this is equivalent to running Steel Plates instead of Iron Plates in a desert biome because there’s more coal, so producing steel is actually cheaper than iron, even though both of them are treated as the same items in the game. You optimize your production lines (codons) to use cheaper materials that does the same thing, to be used in a recipe (protein).
I decided to choose yeast (saccharomyces cerevisiae) as my target. I don’t want to cultivate this dangerous protein for production, only for researching the pathways and underlying systems to design circuitry against it, or to leverage it to create targeted treatments. The reason for yeast rather than E. coli is due to the yeast being a eukaryote, which P. falciparum also is. Having a similar structure will ensure a more similar end result and process.
Anyways, the optimized chain is below
ATG GCT CCA ACC TCT GGG GGA GGT GGA ACA AAA GAT GAA TCT GCA AAA CAC GCC CTT GAC AGT ATT GGT AAG AGG GTT CAT GCC CAG GTG CAA AAT GAA GCG AAG CAA AGG AGT AAT GGG GAT TTA AAG GGT TTA CTT ACA TCT GCA ACT TTA AGT GGT GGT GAA TCT GCT TTT ACT GAA AAC CCT TGT GAG TTG ATT AAG GAA AAA AGA GAA AAA TTA CTT GGT GCT CGT GGC GAA CGT AAT CCG TGT GGG AAT GGT TCT GCT TCT GAG AAA AGA TTT TCA GAA GTG TCT GGG GGG GAA TGC GAT GAC AAA AAG ATT GAA GGA AAC GGA AGA AAT AAC GGT GGA GCG TGT GCA CCA TAC CGT AGA TTA AGT CTA TGC AAC AAA AAT TTC CAG AAA ATC AAT AAT TAT GAC TCC TCT AAG GCA AAA CAT AAT CTT TTG GTC GAT GTC TGT ATG GCT GCT AAT TAC GAG GCC CAA AGT TTA ATT CCT TAT CAC GAT CAA TAC GAT GCA ACA TAC CCA GGA TCA GAC TTT AGC ATG TGT ACA ATG CTA GCT CGT TCC TTC GCC GAT ATA GGT GAT ATT ATC CGT GGT AAA GAC TTA TAT AGA GGT AAA AAG AAA AAA AAA CAG AAC GGC AAA GAA ACT GAA ACT GAA CGT GAT CAG TTG GAG AGC AAA CTT AAG GAA ATA TTT GGA GAC ATA TAC AAT GAA TTG ACT AAC GGC AGA AAT GGA GTG AAG GAC CAT TAC CAG GAT ACA ACA GAT TAC TTT CAA CTA AGA GAA GAT TGG TGG ACA GCA AAT AGA CAT ACA GTT TGG AAA GCC ATC ACT TGT GGC ACT CAT GAA GGT GAC ACA TAT TTC AGA CCA ACG TGC TCC AAT CGT CAG GGG CCT TCC CAG GCG CAC CAC TAC TGC AGA TGC AAC GGT GAT AAG CCC GAC GAT GAT AAA CCC AAT ACA GAT CCA CCT ACC TAC TTT GAT TAT GTA CCT CAA TAC TTA AGG TGG TTC GAA GAA TGG GCA GAA GAT TTC TGT CGT AAG AAA AAG AAA TAC GTC GAT ATC GTG AAA AAG TAT TGT AGA GGC GTT TAT AAT GAT GTT CCT AGA TAT TGT AGT AGA AAT GGT TAC GAT TGC GAA CAA ACG ATC TAC AAG AAG GGA TAT TTT GTT ATT GAT AAA GGT TGC ATC AAC TGT TTG TAT GCA TGT AAT CCT TAC GTT GAA TGG ATT GAG AAA CAG AAA GAA CAA TTT GAT AAG CAA AAA AAA AAA TAT ACC GAC GAG ATT AAT GAA GCT AGT AGG TCT TCC AGG CGT CAG AAA AGA GGA GCT CGT TCA ACT GGC TCT TCC TCT AAC TAC GAC GGA TAC GAG TCC AAA TTT TAC AAT ATC TTG AAA GAT GAT TAT GGA ACC GTT GAT GAT TTC CTT AAA CTT TTA AAT AAG GAA AAA TCC TGC CAA GCA GTC AAG GAT AAT GAC GGT GGC ACA ATA AAC TTT ACA GAG AAA AAT GAC GAT AAA AAC AAT AAC AAC AAA GAT AAG GGT ACA TTT TAC AGG AGT GAA TAC TGT CAG GTC TGT CCA GAC TGC GGC GTC AAA TAT AAC GGT TCA GGC TGG GAG GAG AAA AAA AAG AAC GAC CAA TGC AAT ATT AAG CTA TAC AAA CCG AAA AAA GAT GCC CCT CAT ACT CCT ATA AAG ATT TTG AAG TCA GGA GAG GGA AAA GAG GAA ATA GAA AAA AAG CTG GAG GCA TTT TGT GAT AAA AAG GAT GGT GGA AAC TCT GAT TCC TCC CTG TAC GAT CCT TGG CAA TGT TAT CAA TTT GAT CAA TTG GAA AAG GAC GAG AAG GAA GAG GGT GTA GAT GAC CGT AAC TAT GAT AAC GAT GTT CGT ACT GGA GGT GGC TTG TGT ATC TTA CCA AAT CAA AAG AAA AAC AAA GAG GAA GGT GCT AAC ACA AGT GAA AAA GAT CAC GAC GAA ATA CAA AAA ACA TTC AAT CCA TTC TTT TAC TAC TGG GTG GCG CAC ATG TTG AAG GAC TCT ATA CAC TGG AAA AAG AAA TTG GAA AAA TGT TTA CAA AAT GGT ACG AAG ACC AGA TGC AGA AAT AAT GAA AAA TGT AAT AAG GAA TGC GAG TGT TTC CAG AGA TGG GTA GAG AAA AAA AAA ACG GAA TGG GGA AAA ATA AAA GAT CAC TTT AAA ACT CAA AAT ATC GGT GAT GAA ACA AAC TGC GAT CCT ATT GTA ACC TTA GAA GGT GTT CTT AAG TTA CAA TTT TTG AAC GAG GAT AGT ACT CAA GAT AAA CAG AAT TCA TTA GAT AGT GAA GAG TTG AAA CAT CTA AAG CAC CTT TCC GAA ATG CTG CAG GAA ACA AGT GGT GAC GGC TTA ACA TGC GGC GCA TCA GAT AAT GAA AAA GAA ACT TTG ATG GAC AAG CTA ATC GAA CAC GAA GAG GGT ATC GCA AAG GAA TGT CTT AGG AAG CAA AAT GAA TGT GAA AAA AAA GCG AAA CCA GAA GGG AGG TCT GAT TCA CAC GAC GAC CCG CAG CCA CCA GAC GAT GCC GAT AAC GAG GAT GAT TTA GAT GAT GAA GAC GAT GAA GAC GAA GAG GAA GAA GTA CAG GTC GAA GAT AAT ACT CAA GAA GAG GGC GAA CAA CCG GTC GTT CCA CAA CAA GAA GAA GGT TCC TCT TCC CCG ACT CCT GCG CCT GCT GGT CCT GAT GTG TGC GAT ATC GTA AGC CAG TTA TTC AGT GAC CCT AGT CAA TTT TCA GAT GCT TGT ACA TTG AAA TAC GTA ACT GGG AAG AAT TAT GGC TGG AAG TGT ATA CCA TCT GGT AAC ACA TCT GAC ACC ACA GGT TCT GAA TCA GAA GCA ACT GGA GCT AGA CAA AGG AGG GAT ACA GAC TCT TCT GGT GAC ACG ACA GGA GGT AAA GAT GGT GCG ACC GGT GGT TTA TGT ATA CCT CCG AGG AGA AGA AGA TTG TAC GTG GGT GGG TTA AGT CAG TGG GCA TCT CAG AGA ACA CAA GGA GAA ACT TCT TCC CAG TCC GGT GAA AAC TTG TTA GAG GCA TTT ATT CAG AGC GCT GCC ATT GAA ACA TTT TTC TTA TGG CAC AAA TAT AAG GCT GAA AAT ACT AAA ACT CAA GGC GTT GGA GCA GGT GGT GCC GAT TTC TTA CCT GCT ACT TCT AGT GTT GCC ACG GCT TTA GCT CCT GGC GCT GTT CCA TCT AGG CCA TCT TTG CAA TTG TTG AGT GGT GTT GGT GTC CCT GGG GAA CCA GGT ATG GCC CCG GGT GTC AAG TCT ATC CCT GTA CCG CCA TTA GGT GTA GGT GTT GGT GGA ATT CCC GGT GTT GGA GCA CTA GGC GGC GGG GCA CTA GGT CCA GGT GGT CCA GTG GGT CTG GAT GGC GTA CCA GGT CAA GCA CAG CCG CTA ACT TTA CTT AGA CCT GGC GTC CTG GGC AAC GGA CTG CAG TCT CCT CAA TCA CGT TTA AGA ACT TTA GAT GGT CAT TTC GCC GGT GGT GAA AGT GAA GAT AAA ACC CCT CAA CAA TGG TTA CAA CAA GGT CAT ATT CCT CCA GAT TTC TTA AGG TTG ATG TTC TAT ACA CTT GCA GAT TAT AGA GAT ATC TGT ATT GGT GGT GAT AGA GAT ATA GTA GGT GAC ACG ATA GTT TCC AAC ACA GAA GGT TCA TCC TCT TCC AAG ATA AAA ATT AGT GAG AAA ATA AAA GAA ATA CTG AAT CAC GAT AAC AAA CAG GAG CCG GCT CCA AAG CCT TCT GTT GAA AAG ACC ACC CCG ACT GAA TGG TGG AGT CAA AAC GGT CAG CAT ATT TGG AAC GGT ATG ATT TGT GCT TTA ACT TAC ACG GAT TCA GGT GGA TCA ATT ACA GAA GAC AAG GAT GTT AGA GAC AAG TTG ATA GAT AAA GAT ACC GGC AAG CCT CAA AAA AAC GGG GAT AAT GAT TAC ACT TAT GAG AAA GTT GAA TTG AAG GAC GAC GAA TCA GGT CCT AAG GGT AAT GAC ACA ATT CAG CCA GCT ACT TTA AAG GAT TTC GTT GAA ATA CCT ACA TAC TTC CGT TGG CTT CAC GAA TGG GGC AGC GAT TTC TGT GGA AAA AGG GCT AGG ATG CTG AAA GAC GTT AAA GAT AAC TGT AGG AAT ATT GAC AAA GCC GGC CAT CAT TAT TGT TCC GGC GAC GGT TAT GAT TGT ACC AGA GAC GTT ATC GAA CGT AAT GAT AAG TTC GTC GAT TTG AAT TGT CTA GGT TGC TAC AAG CAG TGT AGA AAG TAT AAG AAG TGG ATC GAC ATT AAG TTC GTT GAA TAC CAT AAC CAA GAA AAA AAA TAC AAG GAC GAA TAT GGG AAG CTA ACA AAA GAC AAA TCT TCT GAT GAT AAG AAG CTA GAG GGC TAT AAA TGT GCT GAA AAT TTC TTG AAG GAA TTA AAG CAC TGT AAG CCG TCC GAA GAC AAC AAT GAT CAG GAT AAT AAA ATA AAC TTT GAC AAG CCT GAA AAG ACA TTT AAT CCA TCT AAA TAT TGT AAG GCT TGT CCT GTC TAC GGT GTT AAA TAT ATA GGG GGA AAT TAT ATA CCG AAT GAA GAA AAG GAC TAT AAG TCA AAG AAG GGT AGG GTC AAA AAG GAG AAC GAT ACC ATC CCC AAG AAC ATA GAA GTC TTA GTG TTG GGT AGA AAG GGG GAA GAG AAA GAT AAG GAC AAA CAT TTG CAT GAC GCT TGC AAA AAC ACG GGT CTA TTT GAA GTA GCT AGA TAT GAA CAA TGG AAC TGC CAA AAG AAA AAA GGA ATT GAT CAA TGT AAA ATC ACT AAA TTT GCT AAC GAT ATA GAT TTC GAC AAA GAT ATC GTT TTT AAT GAA TTC TTT CAA AGA TGG TTA AGA TAC TTT GTC CAA GAC TAT AAT AAG TTA AAG GAT AAA ATA AAT CCC TGT ACT AAG AAA GAA ACG GAA AAG GAA AAA GAA AAA AGT TAT AAA TGT ACC CAA GGT TGC AAT GAC AAA TGC GAA TGT GTC AAG GAA TGG TTG TCA AAA AAA AAG CAG GAA TGG ACT CAA ATT AAG ACA CTG TAC AAA CAG TAC TCA AAA ATA TCT GAT CAA GAA ATA GCC TTT AGG GTA AAA TCA TAC TTC GTT GAC CAA GGA TTA TTT GAT AAT GAC TAT AAA AAA GCA CAA GAG GTC GTC GAG AAA CCC TGT GAC AAA GAG AAA TTG TGG GGT TGT ACC GGC GAT AAT TTA AAG GAA GGA GAA GAT CCA GGT AAA TGT CAC ATG GGT GAT TTT ATT ACT AAT TTA ATC AGC AAA TTG CAA AAG AAA ATT GAT GAC TGT AAT AAG AAT CAA GCC CAG AAC AGC GTT GAG ACA CAA CCA AGT GAT GAA AAT ACC GCA CAG TGC CAA GAT ACT CAT CCA GAT GAT GAA GAG GAT CTT TTG TTG GAA GAA AAC GAA AAT CAG GTC GCT CAG CCT AAT ATT TGT CCT AAT CAA GTT GAA GAC AAG AAA ATA GAA GAA GAA GTG GAA AAA TGT GAA ACC GCA CAA ACG ACC GCG GAA GAG ACA GCC GCA GCC GGC GGA GAG AGA CAG ACT CCA CCT GCC CCT GCG CCA GCG GCA CCA CCC TCC CCT CCT AGA CCA TTA CCA AAG CCG AAA CCC CCA AAA CCT GAC TTG CCA CCT GCC CTT AAA AAT GCT ATG CTT AGT AGT ACG ATA ATG TGG TCT GTG GGC ATT GGG TTT GCT GCA TTT ACA TAT TTT TTT TTG AAA AAG AAA ACG AAG GCT TCA GTA GGT AAC TTG TTT CAA ATT TTA CAG ATT CCA AAG GGA GAT TAT GAC ATT CCG ACA TTG AAG TCA TCT AAT AGA TAC ATT CCC TAC GCA TCT GAC AGA TAC AAG GGT AAA ACA TAT ATC TAT ATG GAA GGT GAT AGC TCC GGA GAT GAG AAG TAT GCT TTT ATG TCC GAC ACC ACT GAC GTC ACT TCT TCA GAA TCT GAA TAC GAA GAA TTA GAT GTT AAC GAC ATC TAC GTG CCC GGT TCT CCT AAA TAT AAA ACG CTG ATT GAA GTC GTT CTA GAG CCA AGT GGT AAT AAC ACT CCC ACA AGT GAC ATA CCT AGT GAC AAC ACA CCA ACC CCC CAG CCC ATA ACC GAC GAC GAG TGG AAT CAA TTG AAG CAC GAC TTC ATC AGC AAT ATG TTG CAA AAT ACG CAA AAT ACC GAA CCT AAT ATC CTG CAT GAC AAT GTA GAT AAC AAC ACC CAT CCG ACT ATG AGC AGG CAC AAC ATG GAT CAG AAA CCA TTC ATA ATG AGC ATC CAT GAC AGA AAT TTG TTT AGT GGC GAA GAA TAC AAT TAT GAC ATG TTC AAT TCC GGT AAC AAC CCA ATT AAC ATA TCT GAC AGC ACC AAT TCA ATG GAT AGT TTG ACC TCT AAC AAT CAT TCT CCC TAC AAT GAC AAA AAC GAT TTA TAC TCA GGA ATT GAC TTG ATA AAC GAC GCA CTG TCA GGG AAT CAC ATT GAC ATT TAT GAC GAG ATG TTA AAA AGA AAG GAA AAC GAG CTT TTT GGT ACA CAA CAC CAC CCT AAA AAT ATA ACA TCT AAT AGA GTT GTG ACT CAA ACG TCT TCT GAT GAT CCT ATT CAT AAT CAA TTG AAC CTG TTC CAC ACA TGG CTG GAT AGA CAC AGA GAT ATG TGC GAA AAG TTA AAA AAT GAT AAC GAA CGT CTA GCG AAA TTG AAA GAG GAG TGG GAA AAT GAA ACC CAC TCT GGA GAC ATA AAT AGT GGT ATT CCT TCC GGG AAT CAT GTA CTA AAT ACT GAT GTC TCA ATT CAA ATC CAT ATG GAC AAC CCT AAA CCA ATT AAC GAG TTT ACC AAC ATG GAT ACG TCA CCT GAC AAA TCT ACA ATG GAT ACT ATA ATT GAC GAC TTA GAA AAG TAC AAT GAA CCA TAC TAC TAC GAT TTT TAC GAA TAT GAT ATA TAT TAC GAT GTC AAT GAT GAT GAT AAA ACC TCT ATG GAT AAC AAC AAT AAT TTA GTA AAC AAA AAT AAT CCT GTG GAT AGC AAT TCA TCT ACT TAC AAC CAT CAT AAC CCA GCT GAT ATT AAC AAA ACT TTC GTT GAT ATT AAT AAC CAC AAC CAA CAT CCA ATT GAA AAA CCC ACA AAA ATA CAG ATT GAA ATG AAT AGC AAC AAT AGA GAA GTG GAT GAG CAA CAA TAT CCT ATT GCC GAT ATA TGG AAT ATT
3.4. Great, we have a sequence. Now what?
Part 4.Prepare a Twist DNA Synthesis Order
4.1. Insert […]
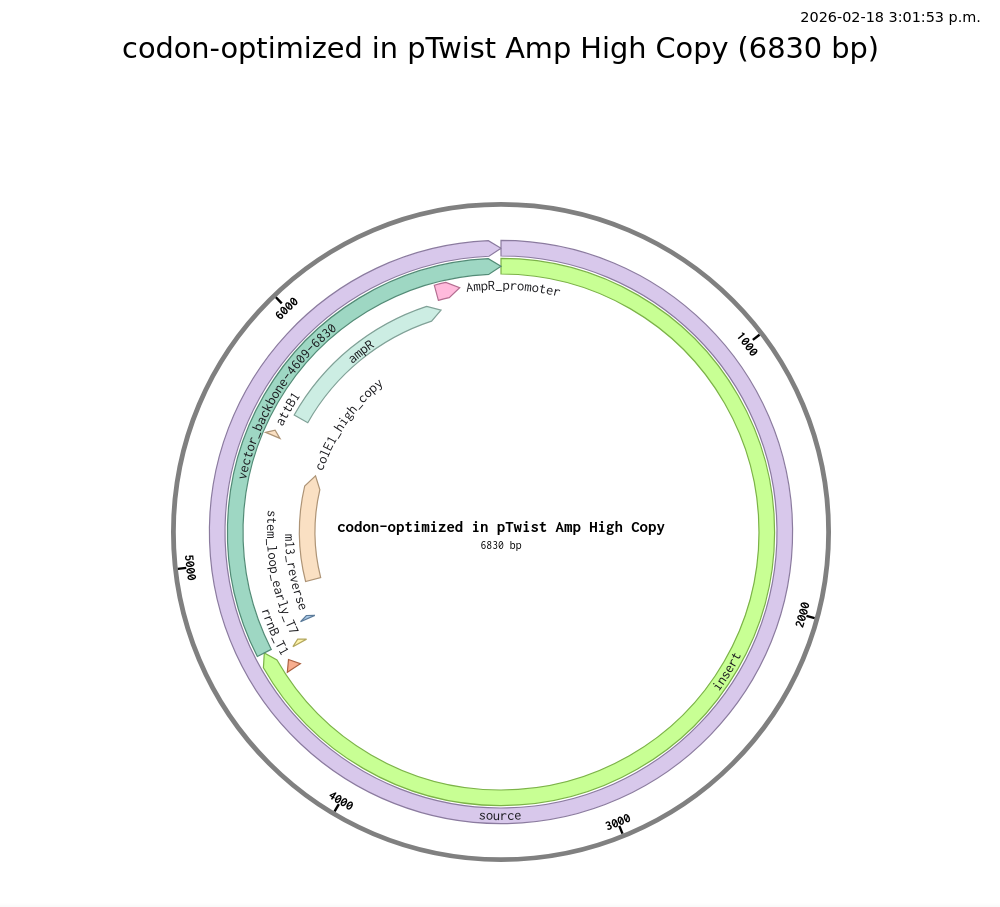
4.2. Plasmid
Funny looking plasmid
Part 5. DNA Read/Write/Edit
5.1. DNA Read
(i) The DNA I am looking to read is one of the proteins of a family called PfEMP1. This protein is one of the many that is non-randomly generated by Plasmodium falciparum to avoid detection from the immune system. I want to study this and similar proteins in this family to see if we can find out what the original pathway is to make personalized treatments for malaria.