A post-mitotic asymmetric condensate disassembly module for autonomous state bifurcation The project is, essentially, about a system that is able to generate one, or very few, condensates, that upon cell division, release their content in the cells that inherited the condensate. This content could be mRNAs of different transcription factors that are able to drive cell-fate differentiation. This proposes to be an step to unlock the potential of autonomous cell-fate differentiation, an essential tool for multicellularity, in synthetic multicellular systems.
A post-mitotic asymmetric condensate disassembly module for autonomous state bifurcation
The project is, essentially, about a system that is able to generate one, or very few, condensates, that upon cell division, release their content in the cells that inherited the condensate. This content could be mRNAs of different transcription factors that are able to drive cell-fate differentiation. This proposes to be an step to unlock the potential of autonomous cell-fate differentiation, an essential tool for multicellularity, in synthetic multicellular systems.
This project is divided in two sub-projects: the post-mitotic activation circuit and the single condensate formation.
A detail description of the project is discused in the following card. Next is the individual final project documentation required.
¿How would it work? ¿What are the components? ¿Why those components?
SECTION 1: ABSTRAC
SECTION 2: PROJECT AIMS
SECTION 3: BACKGROUND
SECTION 4: EXPERIMENTAL DESIGN, TECHNIQUES, TOOLS, AND TECHNOLOGY
SECTION 5: Results & Quantitative Expectations
SECTION 6: ADDITIONAL INFORMATION
Subsections of Individual Final Project
Technical description
General function
Description:
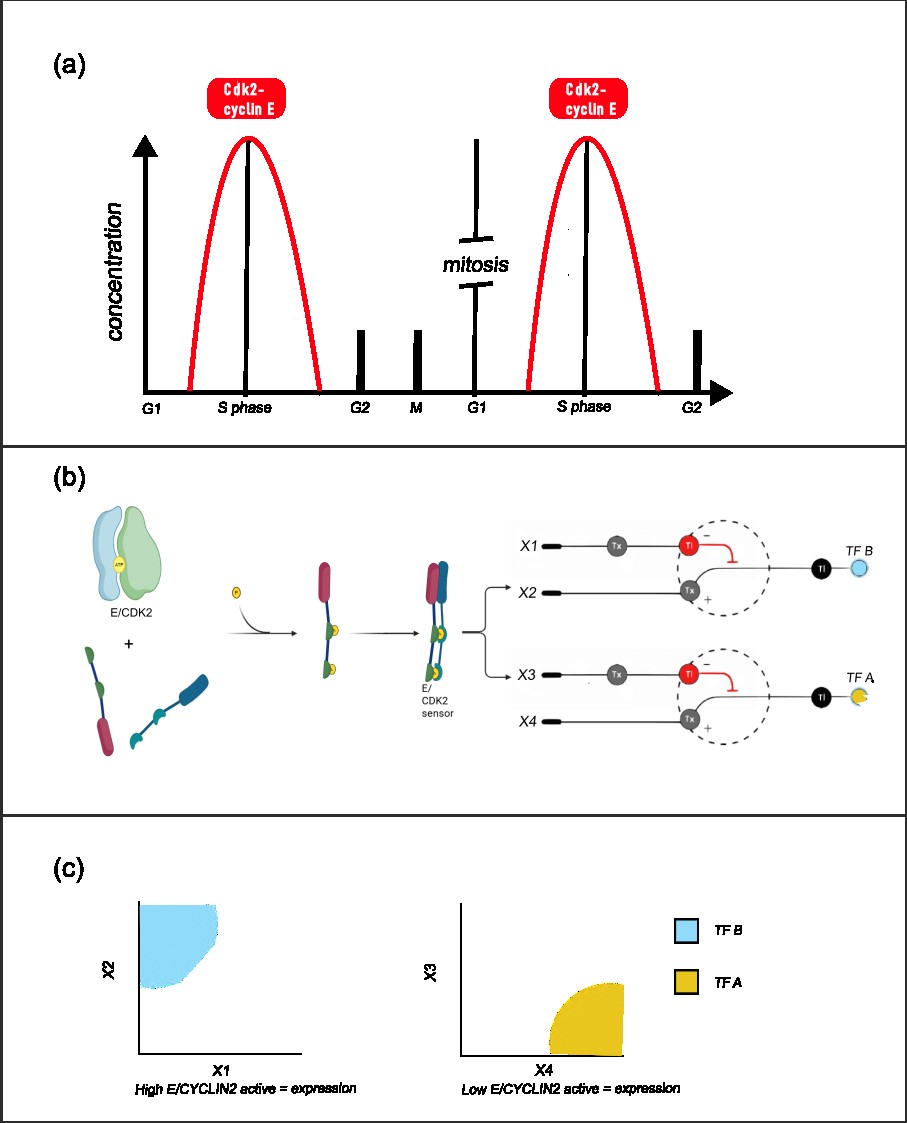
The system achive this by sensing the gradient of the active form of Cdk2–Cyclin E kinase. It sense the high and the low concentrations usign two cellular neural networks-like thresholds circuits [Fig. 1], circuits that put thresholds of activations based on their inputs concentrations, where as outputs i would have two transcriptions factors of short life.
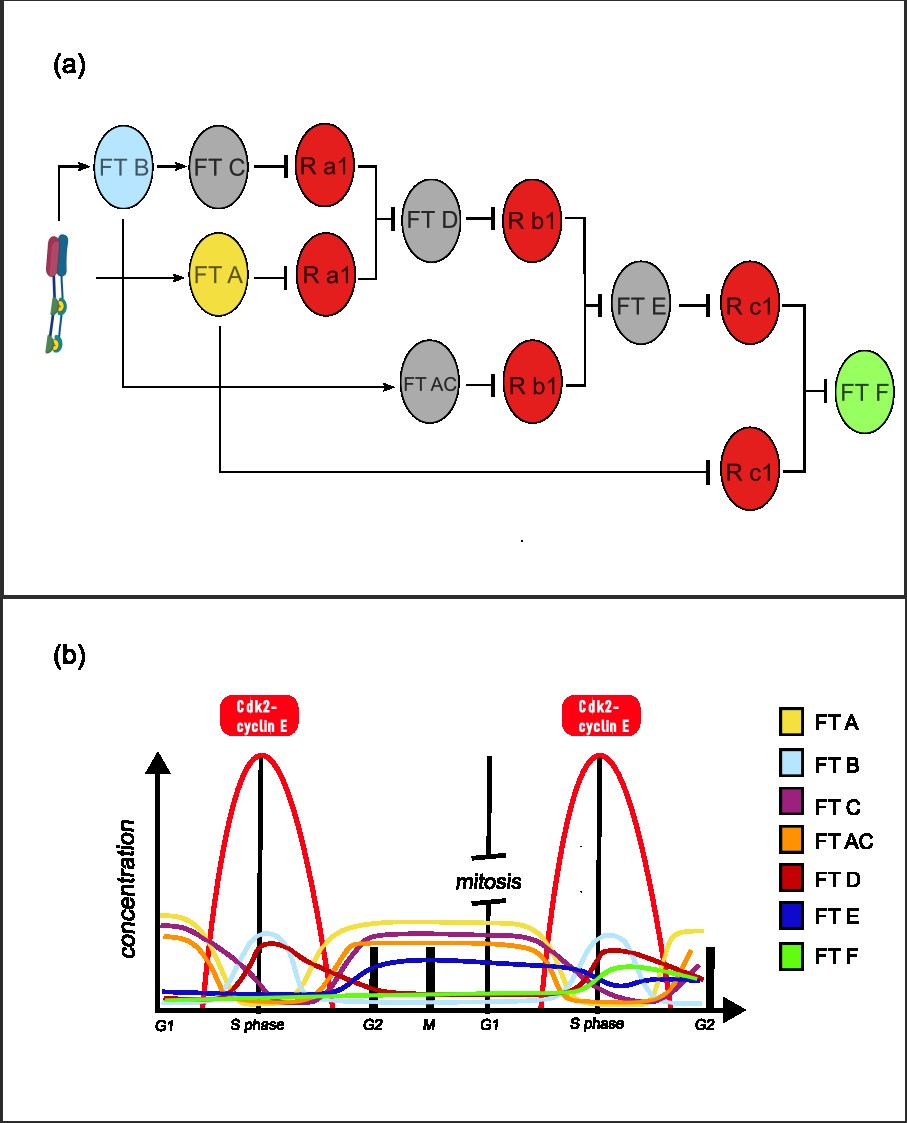
Once the system sense a low concentration of active cdk2-cyclin E kinase, TF A is going to be expressed and it would express TF C (long life). Then, as the active cdk2-cyclin E is growing (high threshold), it would express TF B, wich is going to work with TF C (long life) forming an AND gate to express TF D (long life). Then, when active kinase complex is low, TF A would be active again. TF A also is going to express another short life TF AC, which is going to work with the TF D expressed earlier forming another AND gate, expressing TF E (LF). Finally, TF E is going to form an AND gate with TF B, expressing the final TF F [Fig. 2]. The TF F could express another TF to repress all the components of the system, with a protease, to shootdown the system. The AND gates implemented work by repression of an repressor, where there a two copies of the repressor, each under a promotor with different control sites. The repression of both promoters shutdown its expression and enable the activation of its target.
A pseudo-diagram:
Initial state: low Cdk2 → TF A → TF C, TF AC.
Firts high pulse: TF B + TF C → TF D.
Low state: TF A → TF C, TF AC + TF D → TF E.
Second high pulse: TF B + TF E → TF F.
This archichecture enable having a memory of a past high concentration of a substrate, being able to response different upon that event happen again.
The CNN-like threshold circuits work in the following way:
A Cdk2-cyclin E phosphorylation sensor is designed. This sensor should be specific, have a great dynamic range and have a low basal noise.
This sensor should be a transcription factor, that active upon phosphorylation. This TF ks (kinase sensor) should express two inputs of the CNN (x2 and x3). The first sequestron have as inputs x1 (an RBP that sequest the mRNA of x2) and x2, being TF B, expressed by the TF ks. X2 is going to be active if it suparss x1 quantity, imponing a threshold. As the Cdk2-cyclin E active form is growing, x2 also is going to grown.
The second sequestron have as inputs x3 and x4, where x3 is a RBP expressed by the TF ks, and x4 is TF A. As more TF ks is going active, less TF A is going to be. [Fig 1]
This is just a general description of the system. The specific parts of the system are interchangables and changes on the need. Distintcs transcription factors could be used, distintc tags to mark the half life of the proteins could be incorpored.
In order to exchange the molecule sensed, the sensor is the part to change, having in mind the time dynamics that the another molecule is going to have in the cell, as well as the range of operation of the sensor.
Figure 1: (a) Dynamics of the cdk2-cyclin E complex, with its maximum between G1 and S phase. (b) Function of the E/CDK2 sensor. A chemically inducible dimerization sensor is designed, where two phosphorylation motif, a docking motif and a transcription activator domain is added to one monomer, and a 14-3-3 recognizer motif and a DNA binder domain is added to the second. Upon phosphorylation by E/CDK2, the dimerization induce the expression of its targets genes. This genes are components of the CNNs, where regulation have a downstream effect on (c) Space of expression of the cellular perceptron, where if x1 is low and x2 is high, there would be TF B, and if x3 is low and x4 is high, there would be expression of TF A. As x2 and x3 is inducible by the E/CDK2 sensor, TF A and TF B expression depend on the active Cdk2–Cyclin E complex concentration.
Figure 2: (a) Transcriptional circuit for memory of a high concentration past substrate. Several transcription factors 3 AND gates are implemented, where the repression of two copies a repressor release its target. Repressors for each gate its diagramed as R(a, b, c) and its copy as Rx(1, 2). This circuit enable expression of TF F upon the concentration of Cdk2–Cyclin E active complex goes high again. (b) Concentration of the circuit components over the Cdk2–Cyclin E dynamics.