Individual Final Project
Approaching the challenge of the individual final project 💡
So, here we are. On the page of my individual final project documentation for the 2026 HTGAA course. Before starting to describe and analyze my idea for the final project, I would like to say a few words about approaching this challenge.
Full disclosure: the opportunity to develop a SynBio-related idea into an individual final project has been one of the primary reasons I wanted to attend HTGAA. Whether right or wrong, I thought of the individual final project as a miniature of the iGEM brainstorming and design process that does not necessarily require spending time on a lab bench but that could involve a wet lab component in terms of virtually assembling a construct and generating an experimental plan. In that context, I skimmed through my small notebook of ideas.
I have been passionate about Synthetic Biology for the past six - seven years (ever since I heard about it by accident at a conference), so, during that period, I have been collecting ideas for SynBio projects in a small notebook. Inspiration for those ideas can stem from anything: I might have noticed a beautiful flower on a walk and its shape might have triggered a thought, discovered how a device actually works through a YouTube video and wondered whether there is a biological system that functions in a similar way, or simply read a passage in a literary text or had a conversation with a friend and a concept that I found intriguing popped into my head. In all cases, I wrote down that idea and revisited it at a later time point, for instance, when I browsed my notebook to decide if there were any ideas in there suitable for an HTGAA individual final project.
Several of the ideas I have documented in the notebook are either not SynBio-oriented, too vague, or purely theoretical. However, some of the concepts display the potential of evolving into real Synthetic Biology projects and, I have to say, I did enjoy this small-scale browsing evaluation of them, as it happened in a timely fashion, specifically as I am figuring out the topic I would like to explore for my PhD. But back to the issue at hand, among those ideas, I ended up selecting three for my final project proposals that fulfilled certain criteria (always according to my personal, very subjective point of view): they had to strike a balance between demonstrating a certain degree of creativity and originality, while concurrently being feasible enough to substantially investigate within the duration of a semester-long course such as HTGAA. Additionally, ideas that could have several of the techniques taught during the course incorporated into their experimental design would have an extra advantage.
Based on these criteria, I distinguished the three concepts briefly presented below.
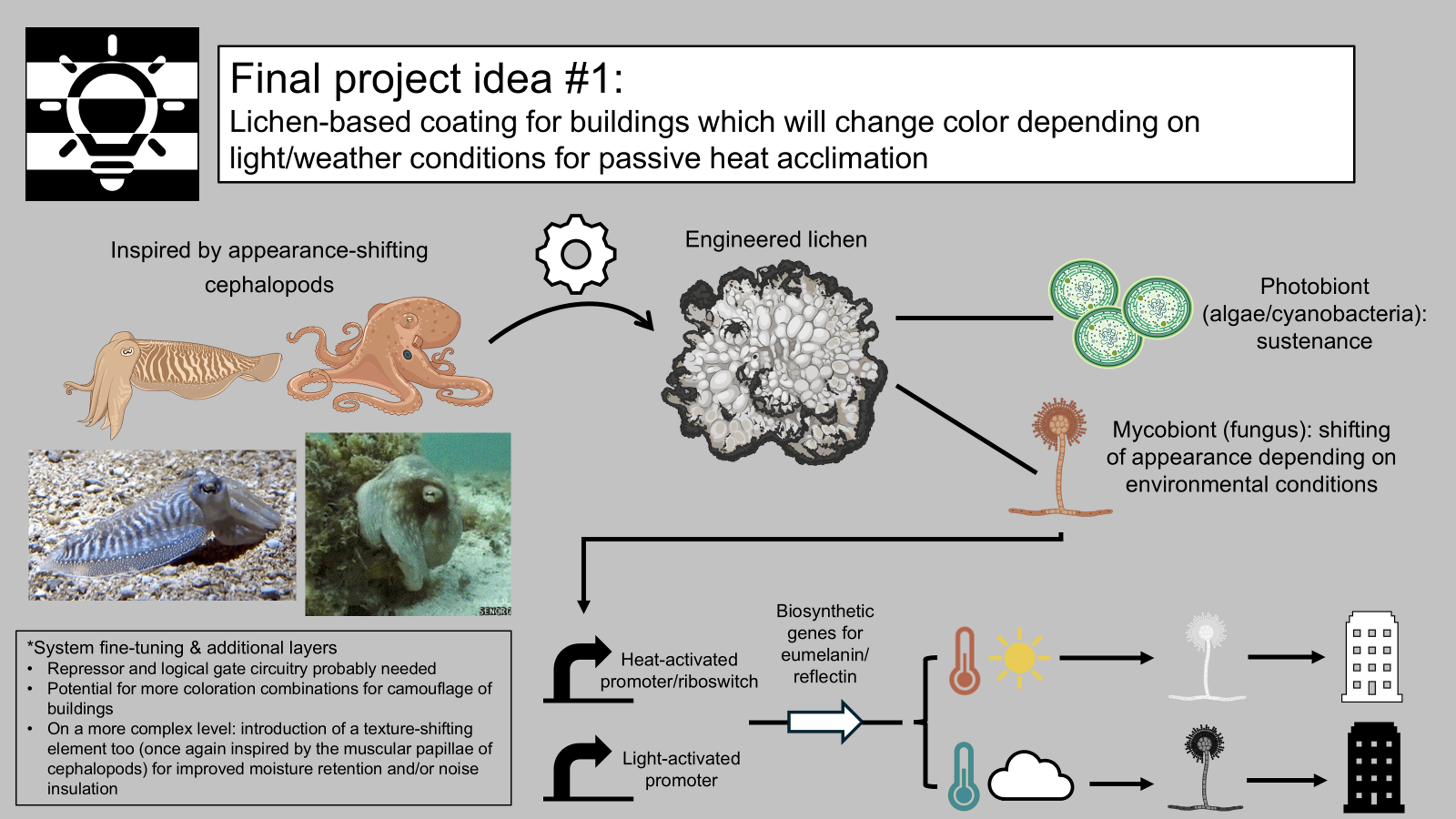
After careful consideration, I decided to proceed with the idea listed first above, namely the cephalopod-inspired color-shifting lichen as a building coating for passive heat acclimation.
Individual final project: Development of a lichen-based building coating capable of changing color depending on environmental conditions for passive heat acclimation ☀️🏢🦑
Section 1: Abstract
It is often overlooked, but our clothes serve as the most intimate, primary layer of shelter, as they function as buffers between ourselves and our surroundings. Among other roles, clothes can be barriers separating the human body from the rest of the world, while providing immediate protection, insulation, and environmental mediation. However, apart from resisting external conditions, clothes can also adapt and harmonize with them as, indeed, we tend to wear darker tones in the winter and lighter colors in the summer to accommodate the lower and higher temperatures respectively, as well as the absence and abundance of sunlight. Based on this rationale and especially now, in a time where climate change and global warming render our need to adapt more imperative than ever, wouldn’t it be interesting to expand this adaptability and pursue harmony between our surroundings and our second and equally fundamental layer of shelter, namely the very buildings we inhabit?
In that context, my individual final project aims at developing a breathing, living biomaterial intended to be used as a building coating. This external layer, just like our clothes, will have the capability of changing its appearance in accordance with its environment by imitating masters of intelligent disguise found in nature, such as octopuses, squids, and cuttlefish. At the basis of this material lies a common dweller of hard surfaces, a lichen, that is genetically modified to respond to its enveloping light and temperature by shifting its color to dark when the weather is cold and cloudy and to white on hot sunny days. These surface-altering properties can be achieved by mainly engineering two genetic cassettes, one with a light-activated and one with a heat-inducible promoter, entangled in a fine-tuned circuit composed of the biological analogues of Boolean logic gates. The genetic circuit’s role encompasses receiving the different environmental inputs and performing “if/then” functions on multiple levels, ultimately generating the most appropriate color output for the given weather conditions, which will, in turn, contribute to the building’s passive and effective acclimation to its surroundings (Figure FP.1).
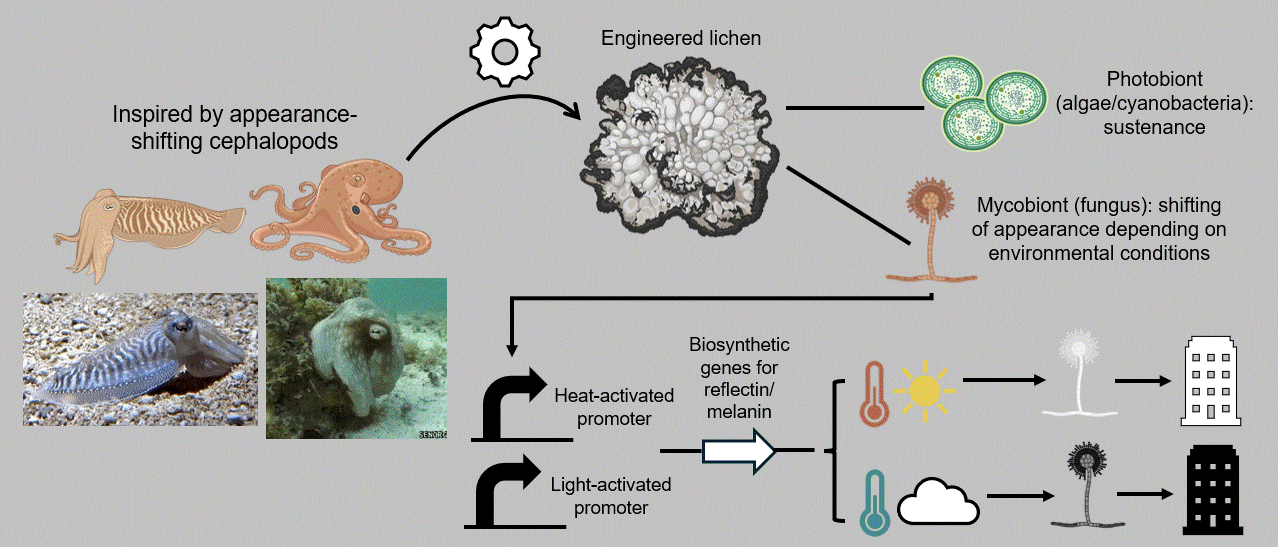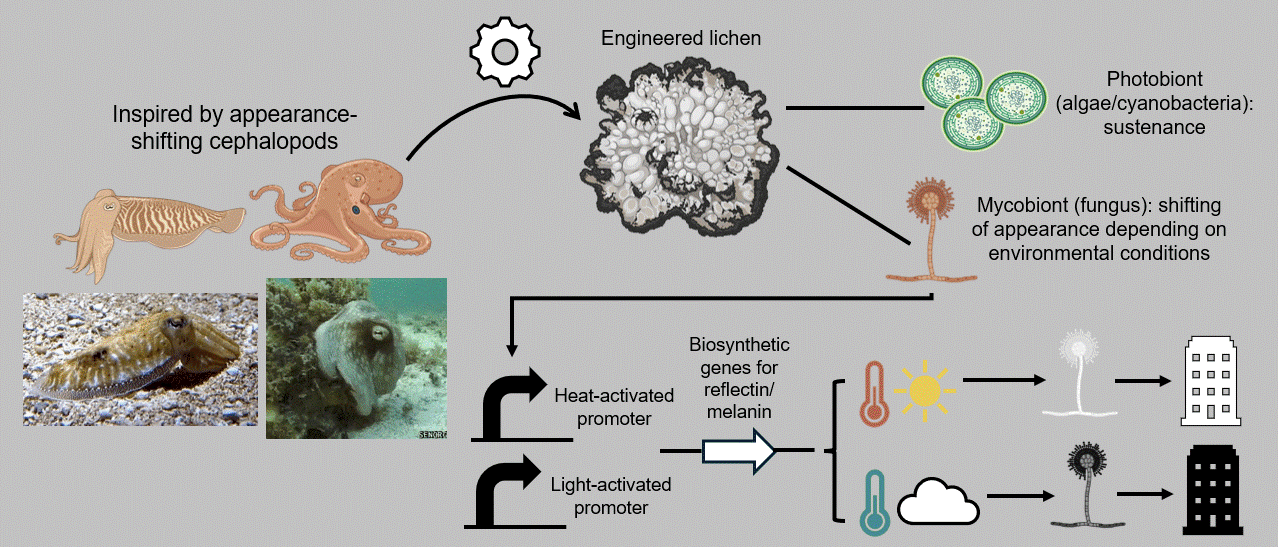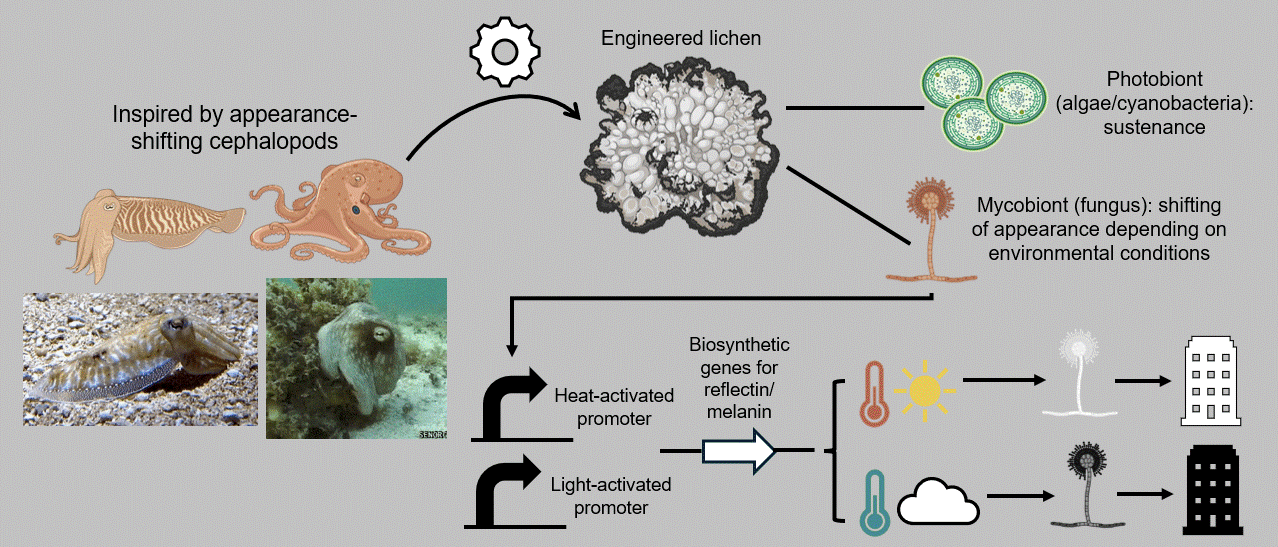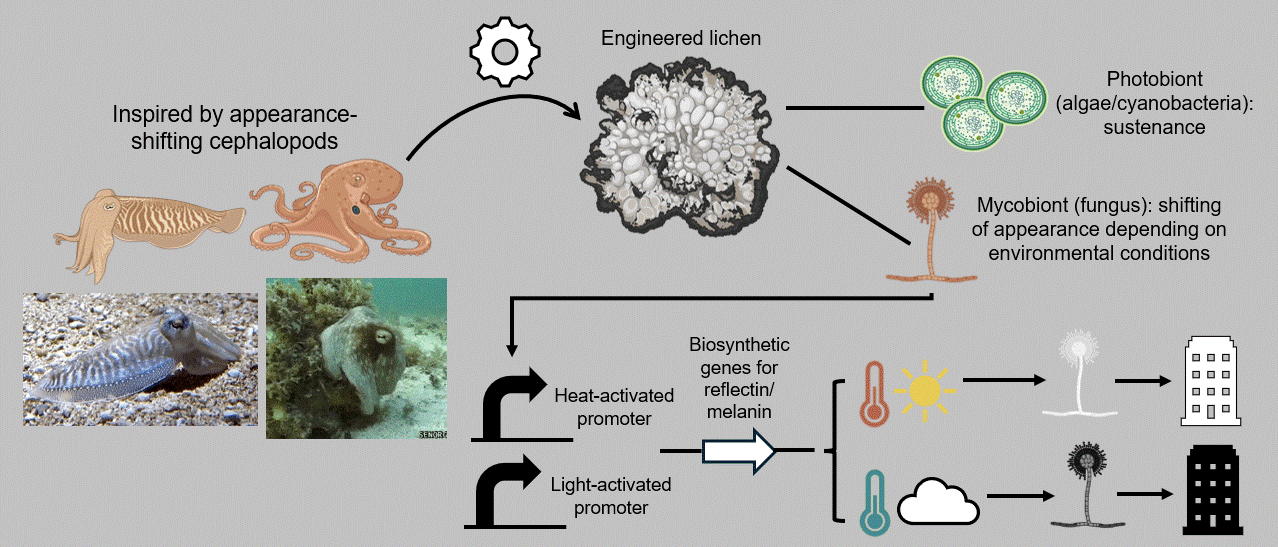 Figure FP.1 Graphical abstract of my individual final project concerning the development of a lichen-based building coating capable of changing color depending on environmental conditions for passive heat acclimation.
Figure FP.1 Graphical abstract of my individual final project concerning the development of a lichen-based building coating capable of changing color depending on environmental conditions for passive heat acclimation.
Section 2: Project aims
Aim 1 (experimental): Simulating the color-changing genetic circuit in Escherichia coli as a proof-of-concept approach
As engineering a lichen is quite a challenging task, for the first aim of my project, I decided to simulate the environmentally responsive color-shifting genetic circuit in a simpler organism, namely the bacterium Escherichia coli. The experimental plan involves adjusting the genetic cassettes for prokaryotic expression and incubating the bacteria under different environmental conditions to induce the production of white or dark coloration. At this point, the output can be measured with several qualitative, semi-quantitative, or quantitative methods.
Aim 2 (developmental): Designing, assembling, and further experimenting with the color-changing genetic cassette in Aspergillus nidulans
The next stage concerns building the actual genetic construct that will be incorporated into the lichen. For the mycobiont member, which, once again, is the part capable of changing color, I have chosen the filamentous fungus Aspergillus nidulans, which, upon engineering, I would like to co-culture with potential photobionts. This step will allow me to investigate the genetically modified fungus’ response in an environment that emulates a lichen thallus. Concurrently, I would like to experiment with novel combinations of myco- and photobionts to identify promising candidates for artificial lichens.
Aim 3 (visionary): Consulting with architects and civil engineers to test a prototype on a structural material
After obtaining a functional engineered lichen, I would like to discuss the opportunity to test a prototype on a structural material with architects and civil engineers. Collaborating with experts specialized in building materials would help optimize the substrate adhesion properties, possibly through a lattice-like installation, to which the lichen can attach and grow, and/or a lichen-supporting scaffold. Alongside this discussion, I also expect that I will need to further optimize the incorporated genetic constructs by performing promoter and transcription factor engineering (among other interventions), as well as genetically modify the selected photobiont too (meaning, in addition to the mycobiont).
Section 3: Background and impact
Buildings that adapt
While I was still a Bachelor’s student, a friend’s friend, who was studying architecture at the time, told me about the first project she had to deliver: designing and making a prototype of a garment. “Why would an architect have to design a garment?” I asked her. “Because our clothes”, she replied, “before buildings, are the first layer we inhabit”. I remember saying “Of course” while being struck by that profound realization. It is true, to some degree, we exist within our clothes but we can also change them depending on our surroundings. We are more inclined to wear thicker, darker garments in the winter and an airy, more breathable, and light-colored attire in the summer. Nevertheless, unlike clothes, buildings cannot really adjust to their environment. Or can they?
There are numerous paradigms of structural interventions contributing to buildings acclimating to their settings. For instance, a prominent architectural feature in the city of Yazd in the desert of central Iran is the wind catcher or “bâdgir” (“بادگیر”) in Persian. Those towers protruding above the rooftops of Yazd (among other cities and towns in the Middle East) provide a natural ventilation system by capturing the incoming cooling wind, circulating it around the dwelling below, and allowing the warmer, less dense air to rise above and escape through another opening, aided by the pressure within the building 1 (Figure FP.2A). Then, skyscrapers hosting vertical gardens, such as the Oasia Hotel Downtown (Figure FP.2B) in Singapore, which is wrapped in a red steel mesh facade supporting more than 20 species of climbing plants 2, receive shading, heat insulation, as well as natural air conditioning thanks to the greenery’s transpiration. Another example of building acclimation is seen in the white houses of the greek islands of Cyclades in the Aegean Sea (Figure FP.2C). The external walls of the buildings are painted white to reflect the incessant sunlight and keep the interior at a comfortable temperature.
 Figure FP.2 Photographs of constructs with elements that help them passively adapt to their surroundings. Figure composed of material from an article in the Tehran Times, a visitor’s experience at Oasia Hotel Downtown, and a personal collection.
Figure FP.2 Photographs of constructs with elements that help them passively adapt to their surroundings. Figure composed of material from an article in the Tehran Times, a visitor’s experience at Oasia Hotel Downtown, and a personal collection.
Being an architecture enthusiast myself, every spring, I visit several buildings of architectural significance in the context of the Open House event in my hometown of Athens, Greece. In 2024, among them was the design and technical office of DECA Architecture, a team of architects based in Athens 3. Their office was an old industrial complex repurposed to house a professional studio and designed to feature structural traits for targeted passive acclimation. I was utterly amazed by the interior of the space, which sported high ceilings to promote air circulation, a Heraklith topping for acoustical absorption (Figure FP.3A), and gravity walls on all sides that harness air’s buoyancy by collecting it when it is lighter, warm, and humid at the top and guiding it through the wall cavity and an installed heat exchanger to the bottom as heavier, cool, and dry air. Two more elements that defined the office space include an in-floor piping network serving as a heat source in the winter and as a heat sink in the summer, as well as my personal favorite: a 50m2 waterwall, dividing the large office space from the public central space. Except for visual and noise separation, the water screen provided humidification and dehumidification, as well as radiant cooling depending on the temperature of the water, which was constantly being recycled 4 (Figure FP.3B).
 Figure FP.3 Photograph and GIF from the DECA Architecture office in Athens. Figure composed of material from Transsolar KlimaEngineering’s website 4.
Figure FP.3 Photograph and GIF from the DECA Architecture office in Athens. Figure composed of material from Transsolar KlimaEngineering’s website 4.
All the above testify that, indeed, like clothes, buildings can act as adaptive layers.
Cephalopods that mimic
Although the architectural interventions presented above indicate that buildings can be designed and erected to achieve the conditioning of their interior, they also reveal a major drawback of most structural traits: they are static. In our era of environmental uncertainty due to global warming and climate change, acclimation mechanisms that are highly dynamic, while exhibiting a low carbon footprint and contributing to energy resources conservation, emerge as a crucial need. Because of this, the capability of dynamic change became a priority in my conceptualization of an environmentally friendly system of passive acclimation for buildings that adapts by responding to an appropriate environmental stimulus. Among the modifications analyzed before, I chose the color of external walls as the concept foundation, since I believe it is the most straightforward and feasible trait to manipulate in the context of a Synthetic Biology-driven acclimation mechanism.
To introduce a biological element and draw inspiration regarding the mechanism’s components, I turned to nature to look for organisms with the capacity of dynamic color changes. Having been mesmerized by marine life since I was a child, my mind went directly to soft-bodied (or coleoid) cephalopods, such as octopuses, cuttlefish, and squids. These invertebrates, having lost the shell that is a typical characteristic of other molluscs, evolved unique adaptations to successfully forage and hide from predators, including their ability to imitate their settings and effectively camouflage through dynamic color- and texture-changing 5 6 (Figure FP.4).
 Figure FP.4 Cephalopods displaying their camouflage and color-shifting capabilities: (A) cuttlefish, (B) octopus, and (C) squid. Figure composed of footage originating from New England Aquarium, a video on camouflage in cephalopods, and underwater footage from Alanya, Turkey.
Figure FP.4 Cephalopods displaying their camouflage and color-shifting capabilities: (A) cuttlefish, (B) octopus, and (C) squid. Figure composed of footage originating from New England Aquarium, a video on camouflage in cephalopods, and underwater footage from Alanya, Turkey.
Coleoid cephalopods can dynamically change their color and skin texture owing to the intricate structure of their body’s surface, which consists of four strata (mainly in octopuses and cuttlefish). The top one contains the dermal papillae, highly specialized skin projections that, acting as muscle hydrostats, can either appear smooth or spiky depending on the animal’s immediate surroundings 7 (Figure FP.5). Similar to the papillae, the chromatophores, in the layer right underneath, can be neurally controlled too, with the help of their surrounding radial muscles. The main volume of each chromatophore organ is occupied by a sac full of yellow, red, or dark brown pigment molecules, which can be expanded or retracted depending on the color the cephalopod wants to mimic (Figure FP.5). At the next stratum lie the iridophores, which reflect light at different wavelengths, thus producing an iridescent effect 5 8, followed by the leucophores right underneath them, which also reflect and scatter ambient light, rendering the skin appear white 8 (Figure FP.5).
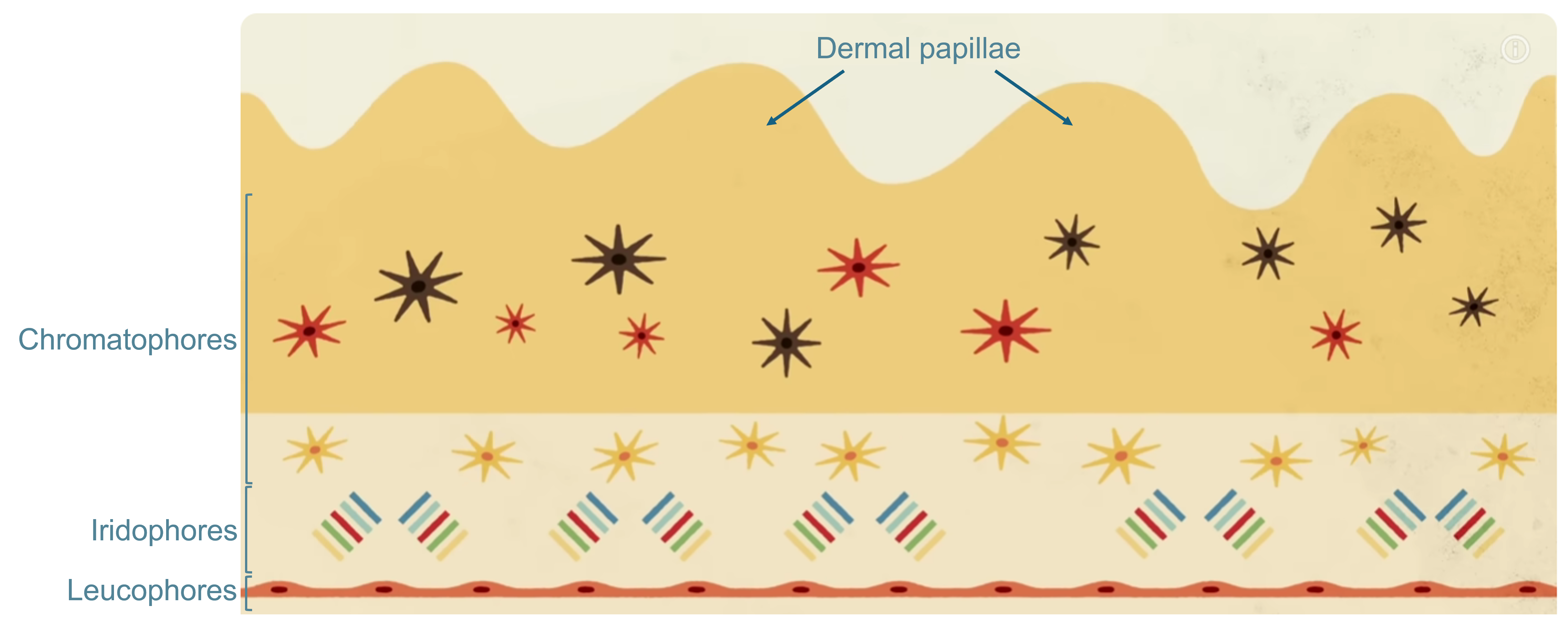 Figure FP.5 Artistic depiction of the distinct layers comprising the skin of octopuses and cuttlefish and contributing to their capacity to imitate their environment. Figure modified from an educational YouTube video on the biology of octopuses.
Figure FP.5 Artistic depiction of the distinct layers comprising the skin of octopuses and cuttlefish and contributing to their capacity to imitate their environment. Figure modified from an educational YouTube video on the biology of octopuses.
Even though cephalopods can apparently generate an entire suite of colors to effectively blend with objects in their vicinity, I decided to limit my options to a more basic palette for my acclimation SynBio application: white, which is highly reflective, and black (dark brown), which is highly absorptive. In octopuses, cuttlefish, and squids, blue-ish white coloration originates from a protein called reflectin 8 9, which forms light-reflecting nanostructures, whereas black/dark brown coloration is derived from ommochrome pigments 9 10.
Materials that grow
After describing the basic mechanism which will serve as a foundation for the color-shifting system I propose, it is also important to analyze the platform in which it will be integrated. For structural applications such as this one, biomaterials derived from mycelium have grown in popularity during recent years due to their aesthetic, acoustic insulation, and fire resistance qualities, as well as for their versatility and biodegradability. Growing “myco-materials”, as they are called, involves propagating fungal hyphae (the branching, threaded shoots of the vegetative bodies of fungi often from the phylum Basidiomycota) into a fibrous lignocellulosic substrate for several days under suitable environmental conditions until they form a composite mass of nearly any desirable shape 11. Among them, one of the most commonly encountered forms are flat, panel-like modules, which can be assembled into a cladding layer positioned on top of an already existing load-bearing skeleton, to reinforce it with properties inherent to mycelium as mentioned before 11.
Despite their plethora of advantages, mycelium-based materials still harbor many drawbacks. Not only do they require strict culturing conditions primarily ensuring maximal darkness and constant access to warm, humid air, they also need to be closely monitored for the emergence of contaminations, therefore, proper sterility constitutes a critical precaution too 11. Additionally, the formulation procedure of myco-structural units includes using a breathable non-cellulose-based material (usually plastic) to prevent the mycelium from permanently adhering to the mold 11. Although they are cultivated from living fungi, another caveat is that most materials derived from mycelium have to undergo a final processing step before being released to the market, involving heat-killing all the living cells 11 12. Myco-composites can, of course, have their color and external pattern altered at previous stages of their fabrication 13, nevertheless, effectively neutralizing the material’s living cells leads to the arrest of all metabolic activity in the fungal hyphae, removing the capability of the material to dynamically modify its appearance, which comprises the main idea of this project.
Thankfully, there have been some recent advances towards maintaining the metabolic capacity of myco-materials and, in fact, in myco-materials that have been engineered to change their coloration once they are exposed to certain conditions too. A study that stands as an example of this reports the development of a “living” material grown from programmable Aspergillus niger mycelial pellets 12. This myco-material is indeed metabolically active and demonstrates a so-called “programmable” color-shifting behaviour, as it has been genetically modified to express genes for melanin biosynthesis upon induction with the monosaccharide xylose (Figure FP.6). Thus, varying concentrations of the inducer result in hues ranging from light grey at lower concentrations and dark brown or even black when xylose is abundant in the material’s proximity 12. However, this membrane can remain alive only for three - four months 12, a period far shorter than the average lifespan of a building, which can exceed 30 years 11. To make matters more challenging, studies like this one are extremely rare in the literature. When I introduced the criterion of the concept being inspired by cephalopods’ appearance-changing capabilities as an additional limiting factor in my research, the selection of materials already devised was even more confined, specifically to a project of constructing a membrane that, powered by the manipulation of its internal air pressure, imitates the expansion and retraction of chromatophores by being inflated and deflated respectively 14.
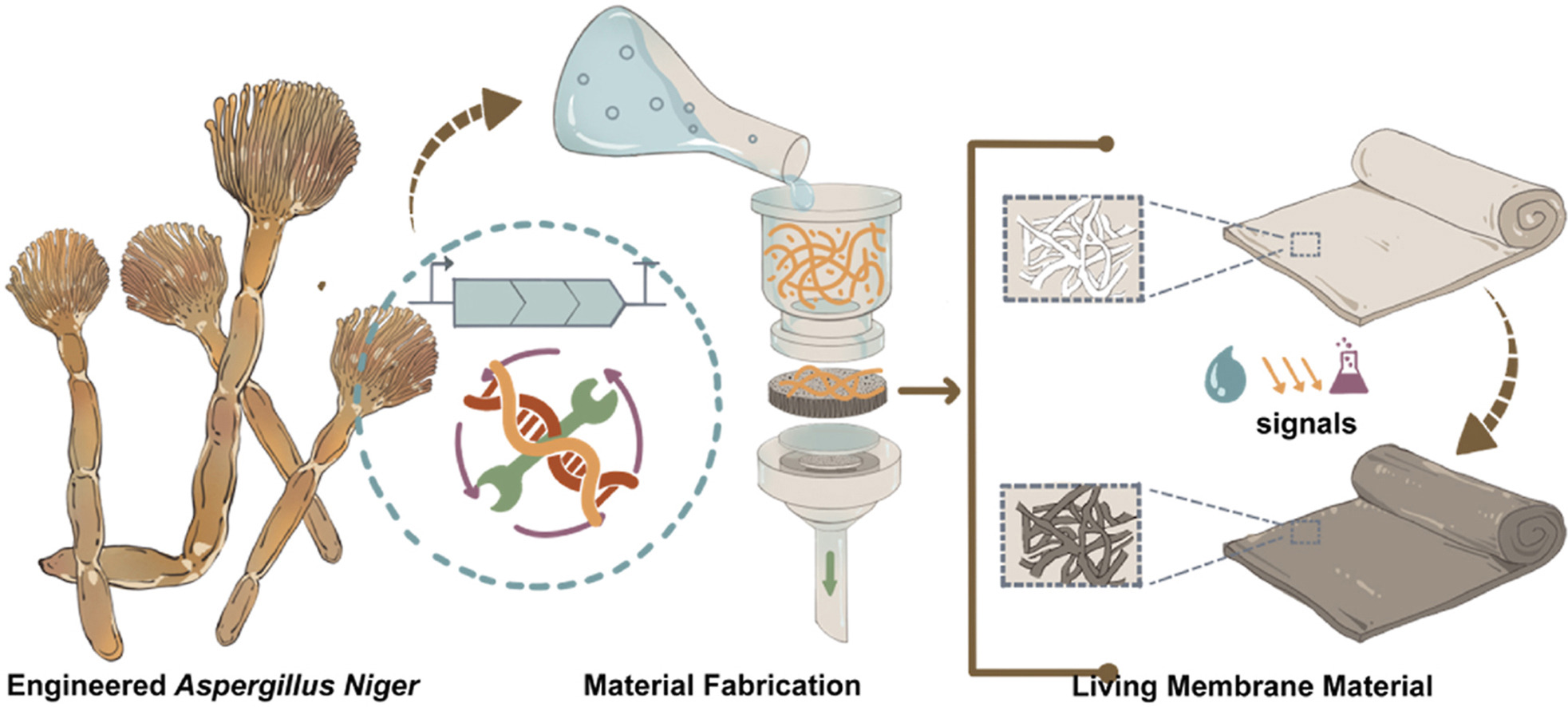 Figure FP.6 Graphical abstract of the paper showcasing the development of a living membrane composed of Aspergillus niger pellets, which have been genetically engineered to respond to varying xylose by altering its coloration from light grey to black. Figure from Li et al., 2023 12.
Figure FP.6 Graphical abstract of the paper showcasing the development of a living membrane composed of Aspergillus niger pellets, which have been genetically engineered to respond to varying xylose by altering its coloration from light grey to black. Figure from Li et al., 2023 12.
The latter study did not involve any Synthetic Biology, so I decided to circle back to biomaterials and myco-structures. At this point of my brainstorming, a material based on a mycelium-like structure, albeit not ideal, seemed to be my best option. What I needed was a fungus-based biomaterial with a particularly long lifespan, so that it does not easily exhaust its color-changing capabilities. Another requirement pertained to the biomaterial being more resilient and less demanding in terms of growing conditions than mycelia-based membranes. Since I mentioned membranes, my conceptualization also emphasized that a biomaterial intended to be utilized as a coating for buildings should, primarily, readily cover many square metres of surface. A truly living myco-material that can naturally stretch over large, flat surfaces then. Once I had listed all the prerequisites, it did not take me long to realize that such an entity already exists: a lichen.
It may seem bizarre, but I will define lichens by firstly writing what they are not: lichens are not individual organisms. They are actually the manifestation of a symbiosis between two or more microorganisms, of which at least one belongs to the fungal kingdom and, of the remaining members, at least one can photosynthesize 15. The fungal member(s) of this mutualistic partnership is known as the “mycobiont”, whereas the photosynthetic part(s) as the “photobiont” 15. The symbionts co-exist in a complex, intertwined configuration called a thallus, consisting of distinct strata inhabited by the fungi, the algae/cyanobacteria, or both (Figure FP.7B) (starting from the surface and moving towards the bottom): the upper cortex, the algal/cyanobacterial layer, the medulla, the lower cortex, and the rhizines (Figure FP.7A) 15.
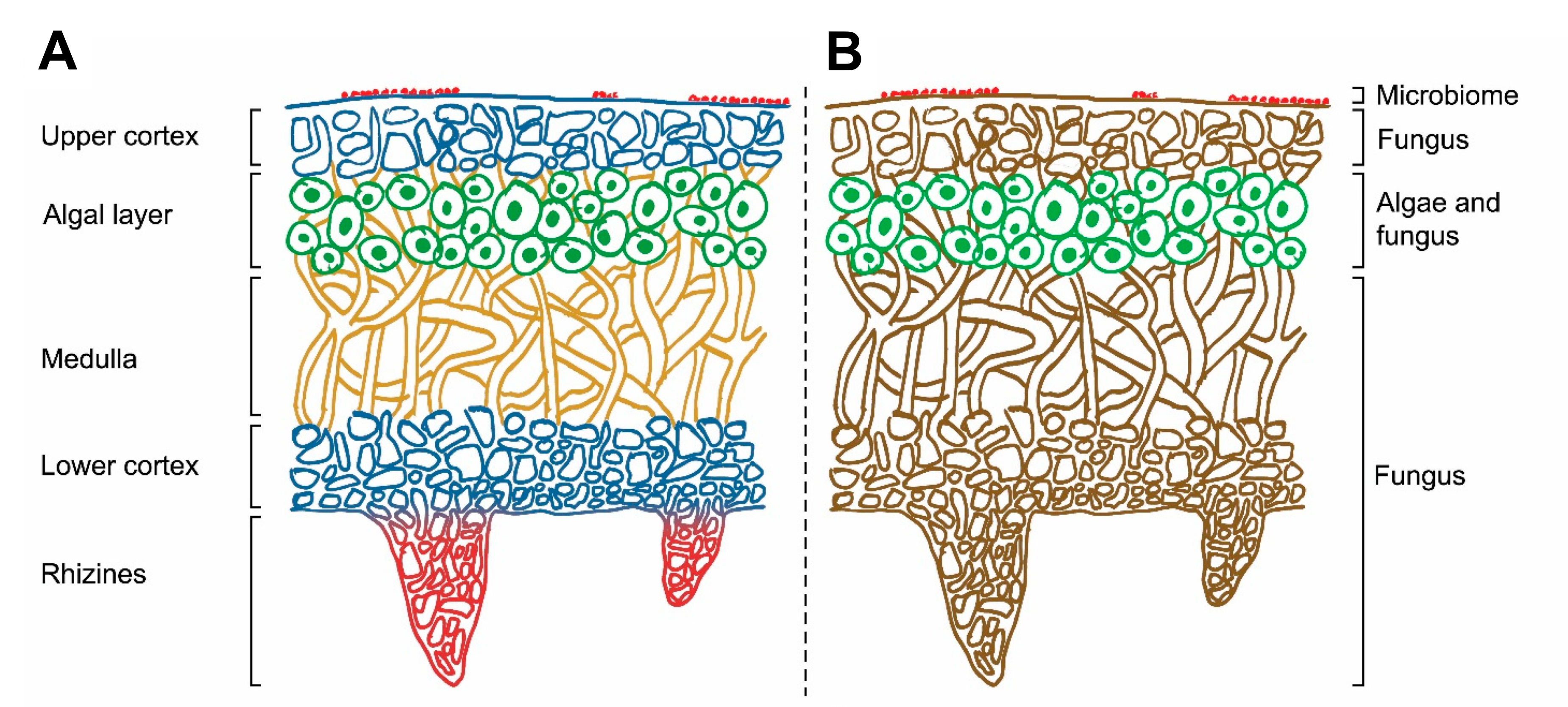 Figure FP.7 Sketch of a lichen’s cross section, displaying its internal architecture and focusing (A) on the different layers of the lichen and (B) on each layer’s inhabitants. Figure modified from Morillas et al., 2022 15.
Figure FP.7 Sketch of a lichen’s cross section, displaying its internal architecture and focusing (A) on the different layers of the lichen and (B) on each layer’s inhabitants. Figure modified from Morillas et al., 2022 15.
Due to the tight symbiotic relationships governing its very existence, a lichen is both more resilient in general and less prone to contamination specifically compared to a material purely composed of fungal hyphae. Furthermore, a lichen is undeniably alive and metabolically active, hence capable of dynamically altering its coloration if programmed to do so. In contrast to myco-materials, which stop developing as well as sequestering carbon during their manufacturing, a synthetic lichen-based membrane can grow alongside a building’s surface, while being self-sustaining thanks to its photobionts. Additionally, those photosynthetic microorganisms procure essential nutrients for the holobiont by fixating atmospheric CO2, therefore dramatically contributing to the lichen acting as a spreading, self-regenerating, functional, and effective carbon sink. Lastly, as organic materials, lichens can be fully biodegraded, while they have evolved to relatively easily colonize hard surfaces without needing extravagant resources or particular environmental conditions. As a personal preference, I found the challenge of genetically engineering a complex life form such as a lichen quite appealing too, while, in my own way, contributing to further fostering lichens as an emerging Synthetic Biology chassis.
This is how the idea of a lichen-based color-shifting building coating was born.
Section 4: Experimental design, techniques, tools, and technology
I decided to divide this section into two parts: the first one concerns a more theoretical design for fungal expression and the second one relates to my first-aim, bacterial design for a genetic circuit.
Fungal constructs
As I mentioned in the Background section, a lichen, and in turn, a lichen-based biomaterial, consists of its mycobiont and its photobiont. For my final project, I have envisioned the two partners of the lichen holobiont to fulfil two separate purposes: the photosynthetic organism’s function will be to autotrophically sustain the actual color-shifting symbiont, namely the fungal partner. Therefore, at this initial phase of the project, it is basically the fungal member of the lichen-based biomaterial that is to be genetically engineered. At this point, you might be wondering why I chose to design the fungal cassettes as well, given that my experimental aim for the course involves implementing the basic idea solely in E. coli, which serves as a simpler prokaryotic expression system. My reasoning behind this was that, in order to effectively simulate the color-changing fungal genetic circuit into a bacterial expression platform, I should first obtain a fairly good grasp of how this final circuit will be structured and what it will look like, hence my prioritizing designing the fungal expression cassettes.
As a first step in this process, I had to choose the organism that would become the fungal partner in my lichen-based coating, so as to select the appropriate genetic parts and, in general, properly tailor the genetic circuit. To this end, I decided to proceed with Aspergillus nidulans, a filamentous fungus species from the phylum Ascomycota. It belongs to the genus Aspergillus, one of the most thoroughly studied genera of filamentous fungi 16. I opted for A. nidulans in particular, as it is a model organism for filamentous fungi, thus many genetic tools have already been established for the species 16 17 18. Albeit not a naturally lichen-forming fungus, A. nidulans can be frequently encountered in synthetic lichens or artificial symbiotic communities built in the laboratory, rendering it a promising candidate for a lichen-based biomaterial 19 20.
Several filamentous fungi, including numerous species of the genus Aspergillus, possess an innate mechanism for the production of darker pigments, especially melanins, once they are exposed to intense light and ultraviolet radiation, which fortifies them against damages in their DNA potentially caused by these factors 21. A. nidulans employs the L-DOPA (L-3,4-dihydroxyphenylalanine) metabolic pathway for melanin production 22, which, if left unregulated, can interfere with the mechanism generating the desired color pattern in the lichen-based building coating, as it does not follow the same coloration-changing principles. This is also the reason I avoided manipulating the genome of other Aspergillus species, such as A. niger, which inherently demonstrates a much higher propensity towards the accumulation of dark pigment molecules. In any case, instead of uprooting the genetic cassette responsible for this response from the fungus’ genome, I wanted to “reprogram” it so that I can use it to my advantage without having to engineer an additional circuit, for example, to induce the production of ommochrome pigments as in cephalopods. For this, I designed a version of the MelC gene, the gene associated with the first biosynthetic step in the pathway towards melanin production in A. nidulans 23, whose regulation can be integrated in the overall environmental control of the appearance-changing genetic cassette (Figure FP.8, left). My goal is to replace the wild-type gene with the regulated version through homologous recombination, so that the fungus retains the capacity to manifest a dark brown/black coloration, although, from this point onward, to do so as dictated by the environmentally responsive cassette for the alteration of its color (Figure FP.8, left).
The version I designed contains two homology arms flanking the MelC gene of interest, along with two additional modules. The first module is a brief DNA sequence, which, upon transcription, is folded into a stem-loop in the nascent RNA molecule (Figure FP.8). This secondary structure is recognized and susequently sequestered by the CasE endoribonuclease 24, which is encoded in another part of the larger color-changing genetic circuit (Figure FP.9). The CasE endoribonuclease is under the control of a heat-inducible promoter, therefore, when the modified fungus is exposed to sunlight, given that the temperature is high, it can “defy its nature” and suppress the synthesis of dark pigments. The second module of this cassette acts as a selection marker for the fungal strains that have been modified to incorporate the “controllable” version of MelC. It is basically an aptamer from the Mango family that strongly emits fluorescence once it binds thiazole orange derivatives 25 26 (Figure FP.8, orange sphere), comprising a reliable and efficient method to identify the “positively transformed” fungal strains (Figure FP.8, right) by incubating all the transformants with the substrate before screening them with flow cytometry.
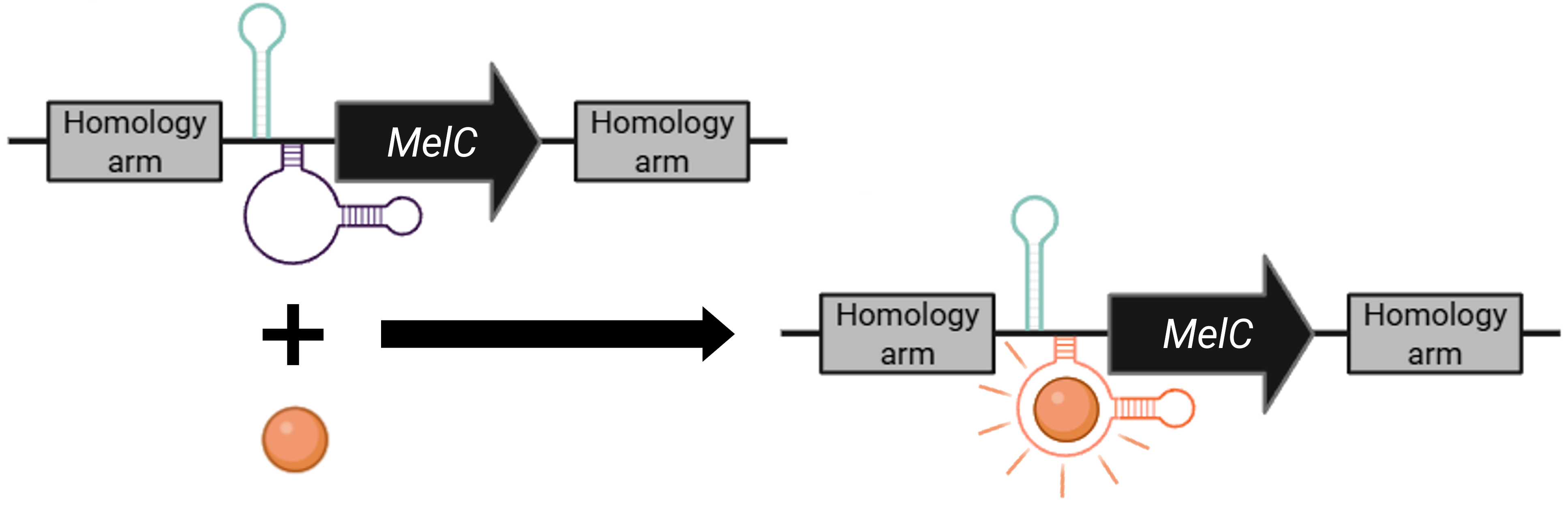 Figure FP.8 Genetic cassette for selection of the A. nidulans strains that have integrated the “controllable” version of the gene enabling melanin biosynthesis. Figure created in BioRender.
Figure FP.8 Genetic cassette for selection of the A. nidulans strains that have integrated the “controllable” version of the gene enabling melanin biosynthesis. Figure created in BioRender.
Now, onto the main circuit, which is designed to determine the color of the lichen-based biomaterial depending on its surrounding conditions. As I mentioned earlier, the expression of the CasE endoribonuclease is regulated by a heat-activated promoter (Figure FP.9, cassette at the top). The promoter is originally from the gene for heat-shock protein 30 (hsp30) from a closely related species, specifically Aspergillus oryzae, whose expression is highly induced at temperatures above 30 - 35°C 27. The primary coding sequence under the control of P hsp30 is, of course, REF8. This gene codes for a reflectin protein from the common cuttlefish, Sepia officinalis 28, and, once expressed, will give the fungal cells a white, reflective coloration at higher temperatures. The two genes in this cassette, REF8 and CasE, are fused with the 2A peptide (Figure FP.9, cassette at the top), a short DNA sequence which corresponds to a peptide spanning about 20 amino acids with an underlying “stop-carry on” translational mechanism. Briefly, when the sequence coding for the 2A peptide is placed between two genes for co-expression, a long, polycistronic mRNA is formed during transcription, which is used as a template for ribosome-mediated translation. When the ribosome encounters the 2A sequence, it skips the formation of a peptide bond between the last two amino acids but continues translation, ultimately enabling the co-translation of equimolar amounts of different proteins from a single transcript 29 30. Another element worth mentioning is the Kozak sequence (Figure FP.9), which, in very broad terms, could be viewed as the eukaryotic equivalent of the Shine-Dalgarno ribosome-binding site in prokaryotes, as it provides optimal start codon context and guides the ribosome to correctly initiate protein synthesis 31. For this initial phase of designing the fungal circuit, I have used the same Kozak consensus in both cassettes (Figure FP.9).
On the contrary, I refrained from using the same terminator in the two cassettes and employed two different ones to diminish the likelihood of unwanted homologous recombination events. The two terminators depicted in Figure FP.9, namely the terminating sequences from genes amyB for α-amylase and trpC for tryptophan synthase, are native to the Aspergillus genus to ensure effective termination of transcription for the cassettes. Focusing on the second cassette (Figure FP.9, cassette at the bottom), since I impaired the light-exposure defence mechanism of the fungus by replacing its main biosynthetic gene for melanin production with one regulated by the color-changing apparatus, I placed the Dsup gene under the control of P ccgA (for “clock-controlled gene”), a fungal promoter activated by ambient white light 32 33. Coupling Dsup, which is a protein discovered to mitigate DNA damage in tardigrades 34, with the light-inducible promoter of the circuit is my proposal to partially restore the fungus’ ability to fortify itself from ultraviolet radiation through an alternative route. Lastly, to integrate the circuit into A. nidulans’ genome, I opted for homology arms that will direct the insert into the pyrG locus (Figure FP.9), encoding orotidine-5′-monophosphate decarboxylase, which is critical for uridine/uracil synthesis 35. For pyrG to serve as an effective selection marker, the original strain that will be utilized for the incorporation of both the “programmable” melanin biosynthesis cassette and the color-changing circuit has to be auxotrophic for uridine/uracil. As the successful recombination of the appearance-altering insert will restore the gene’s function, the selection of modified transformants can be readily performed on media lacking uridine/uracil.
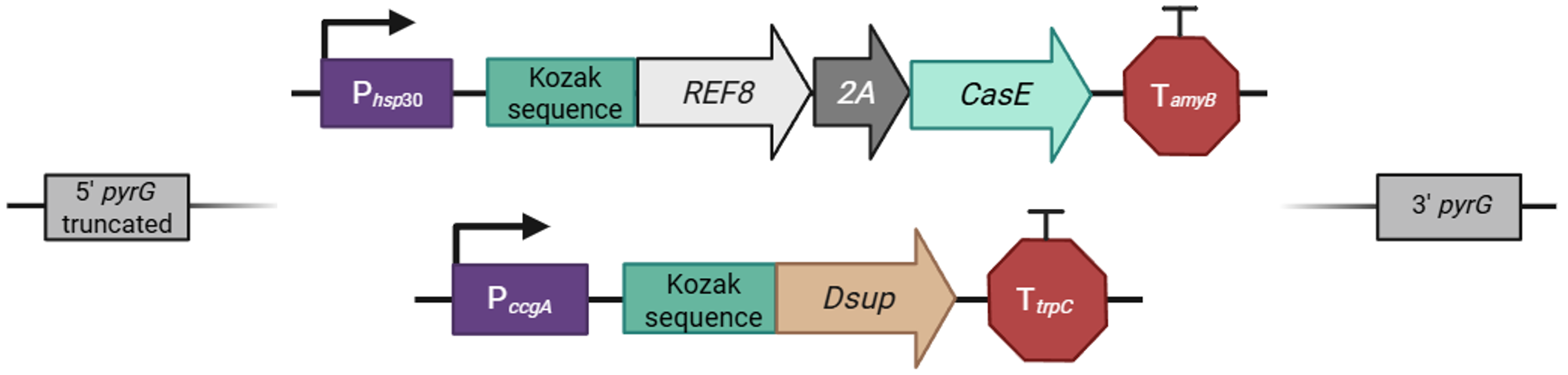 Figure FP.9 Genetic cassettes allowing heat- and light-inducible color alterations of A. nidulans’ hyphae. Figure created in BioRender.
Figure FP.9 Genetic cassettes allowing heat- and light-inducible color alterations of A. nidulans’ hyphae. Figure created in BioRender.
Bacterial constructs
Surprisingly, the genetic construct that would simulate the color-changing effect in bacterial cells proved more challenging to design than the actual fungal construct (Figure FP.9). Among other differences that I will analyze in the following paragraphs, E. coli does not inherently produce melanin, which meant that I had to add a couple of biosynthetic steps so that I could claim I have devised a bacterial genetic circuit that behaves as similarly as possible to its fungal archetype.
Let me begin with the very basics. All the regulatory parts of the original circuit, namely the promoters, the terminators, and the translation initiation sequences, have been replaced with their bacterial counterparts, as fundamental molecular biology systems operate differently in eukaryotes than in prokaryotes. I will certainly explain my promoter selection further, but first I would like to address my RBS and terminator choice. In order to simplify the design, I decided to incorporate the RBS workhorse of bacterial Synthetic Biology in all the transcriptional units, namely the Elowitz RBS (BioBrick entry: BBa_B0034) 36, as it is reliable and efficient. Since bacteria can readily express multicistronic genes, I added the Elowitz RBS multiple times in every transcriptional unit, before each distinct coding sequence (Figure FP.10), for smoother and more robust translation of the proteins contributing to the different colorations. With the same reasoning, I employed the double BBa_B0015 terminator in all genes of the circuit.
Regarding the promoters, it was crucial to include a heat-inducible, as well as a light-activated promoter in the cassettes responsible for the synthesis of reflectin and melanin, to generate the light- and dark-colorations respectively. For the former, I chose P dnaK, one of the most well-characterized bacterial promoters activated by heat. In E. coli, it regulates the expression of dnaK, a protein-folding chaperone triggered as part of the heat-shock response 37. However, in this circuit, I placed REF8, the same gene for reflectin biosynthesis as in the fungal construct, albeit codon-optimized for bacterial expression and with an additional 6x-His tag in its C-terminus for later downstream quantification or visualization (Figure FP.10, first cassette from the top). I linked REF8’s expression with the expression of a dCas9 protein for tight regulation of other components of the construct, on which I will elaborate further down. Back to the promoters, for the latter, the light-inducible one, I proceeded with the P BAD promoter (Figure FP.10, second cassette from the top), which, typically, initiates transcription in the presence of L-arabinose. In an intriguing turn of events, the P BAD promoter can be converted into a light-activated one by implementing the BLADE system (Blue Light-inducible AraC Dimers in Escherichia coli) 38. This system recruits a chimeric version of the AraC operator-binding factor, fused with the blue light-mobilized domain of VVD (Vivid) protein 38 (with blue light being, of course, one of the contained wavelengths in white light). Thus, for this mechanism to function properly and the promoter to be triggered by light, I have also included the VVD-AraC(DBD) (DNA-binding domain) gene in the larger construct, under the control of a separate constitutive promoter (Figure FP.10, cassette to the bottom left). The light-inducible cassette controls the expression of MelC2 and MelC1, two pivotal genes for the biosynthesis of L-DOPA melanin in bacteria that display distant homology to the fungal MelC previously mentioned. This prevalent metabolic pathway for L-DOPA melanin production in bacteria entails the processing of tyrosine, hence the insertion of MelC2, which encodes a Cu-dependent tyrosinase, and MelC1, coding for a protein that carries Cu2+ ions to the active site of MelC2 39, into the bacterial cassette for melanin synthesis (Figure FP.10, second cassette from the top). MelC2 has been tagged in its C-terminus for the same reason as REF8, however, with a different tagging sequence, more specifically a FLAG tag, to distinguish it from reflectin.
Lastly, a few comments about the remaining cassettes of the overall construct, which have been included to fine-tune its color-modifying function. All the remaining cassettes are regulated by two different constitutive promoters, the synthetic P conII 40 41 and promoter J23102 from another bacterial Synthetic Biology fan favorite, the Anderson Promoter Collection. As you may have already noticed, there is a second cassette harboring the genes for melanin synthesis (Figure FP.10, second cassette from the bottom). For the construct to produce the expected color as an output under different combinations of environmental conditions, it was necessary to duplicate the L-DOPA melanin biosynthetic genes and place them under the complementary regulation of two different promoters, the light-inducible one and the constitutive J23102. Nevertheless, J23102, in this specific context, should not be regarded as an entirely constitutive promoter, as it is, in fact, regulated by the expression of the dCas9 protein, which is, in turn, controlled by P dnaK (Figure FP.10, first cassette from the top). dCas9 belongs to the family of CRISPR-associated proteins, however, it has had its catalytic nuclease domain deactivated 42 43 44. As a result, when the heat-inducible promoter initiates its expression, dCas9 forms a complex with the constitutively synthesized gRNA, which was designed for a sequence within the J23102 promoter partially following a protocol published by the Church Lab (Figure FP.10, cassette to the bottom right). The gRNA helps the complex navigate to J23102, thus engaging the promoter, hindering transcription of the downstream genes, and, with this, concluding the design of the bacterial construct.
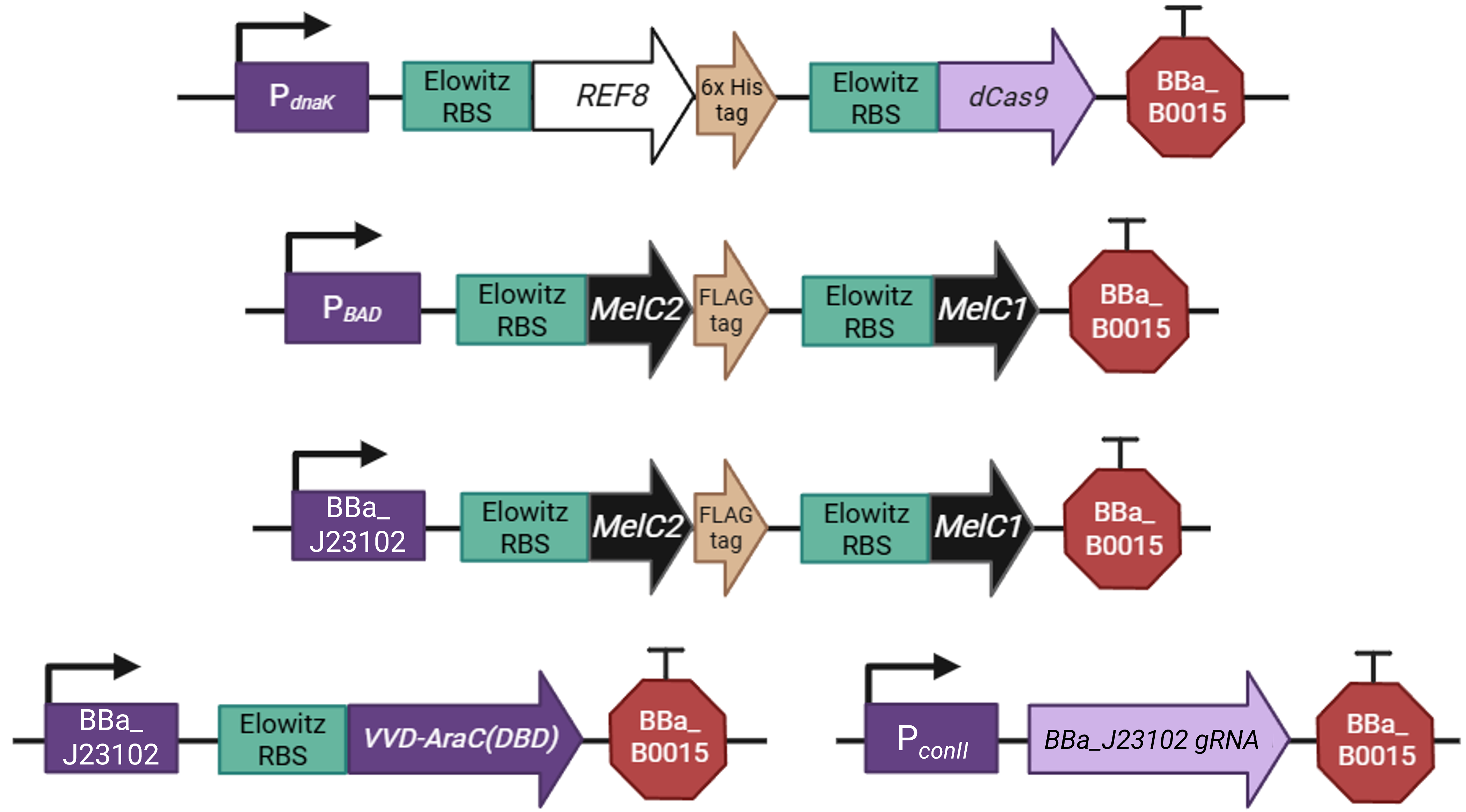 Figure FP.10 Genetic construct simulating the heat- and light-inducible color-shifting effect designed for A. nidulans in E. coli as a proof-of-concept strategy. Figure created in BioRender.
Figure FP.10 Genetic construct simulating the heat- and light-inducible color-shifting effect designed for A. nidulans in E. coli as a proof-of-concept strategy. Figure created in BioRender.
To assemble the construct into a plasmid, I decided to implement the Golden Gate Assembly technique, which, given the number of parts I have to stitch together, seems to be the most suitable approach. As a first step, I assembled the five transcriptional units depicted in Figure FP.10 from their individual parts into four Golden Gate Assembly-compatible level 1 vectors (pTU1-lacZ A - D) derived from the EcoFlex MoClo kit 45 by using the type IIS restriction enzyme BsaI. Due to the design of their EcoFlex backbones, the now assembled genetic cassettes contained in the four level 1 vectors (Figure FP.11A - D) can then be sequentially joined together into the level 2 pTU2-A-RFP backbone from the same kit 45. This second round of Golden Gate cloning involved the utilization of a different type IIS endonuclease, namely BsmBI, to chain all five genetic cassettes together into one plasmid according to Moore et al., 2016 45. This final construct, termed “pCICADA” (Cephalopod-Inspired Color-Altering DNA Apparatus) (Figure FP.11E), harbors the complete bacterial color-changing circuit.
Prior to both level 1 and level 2 assemblies, all the genetic parts were codon-optimized for expression in E. coli and had their sequences modified to ensure there were no BsaI or BsmBI recognition sites in the cassettes. The level 2 backbone I used contains a high-copy origin for replication for robust and efficient plasmid replication. Lastly, I have listed the sources for all the sequences I used for my constructs in a comprehensive table below (Table FP.1), while the full sequences of all the genetic parts I incorporated in the color-shifting circuit, along with the employed construct backbones and assembled plasmids, are stored in my individual final project-dedicated Benchling folder.
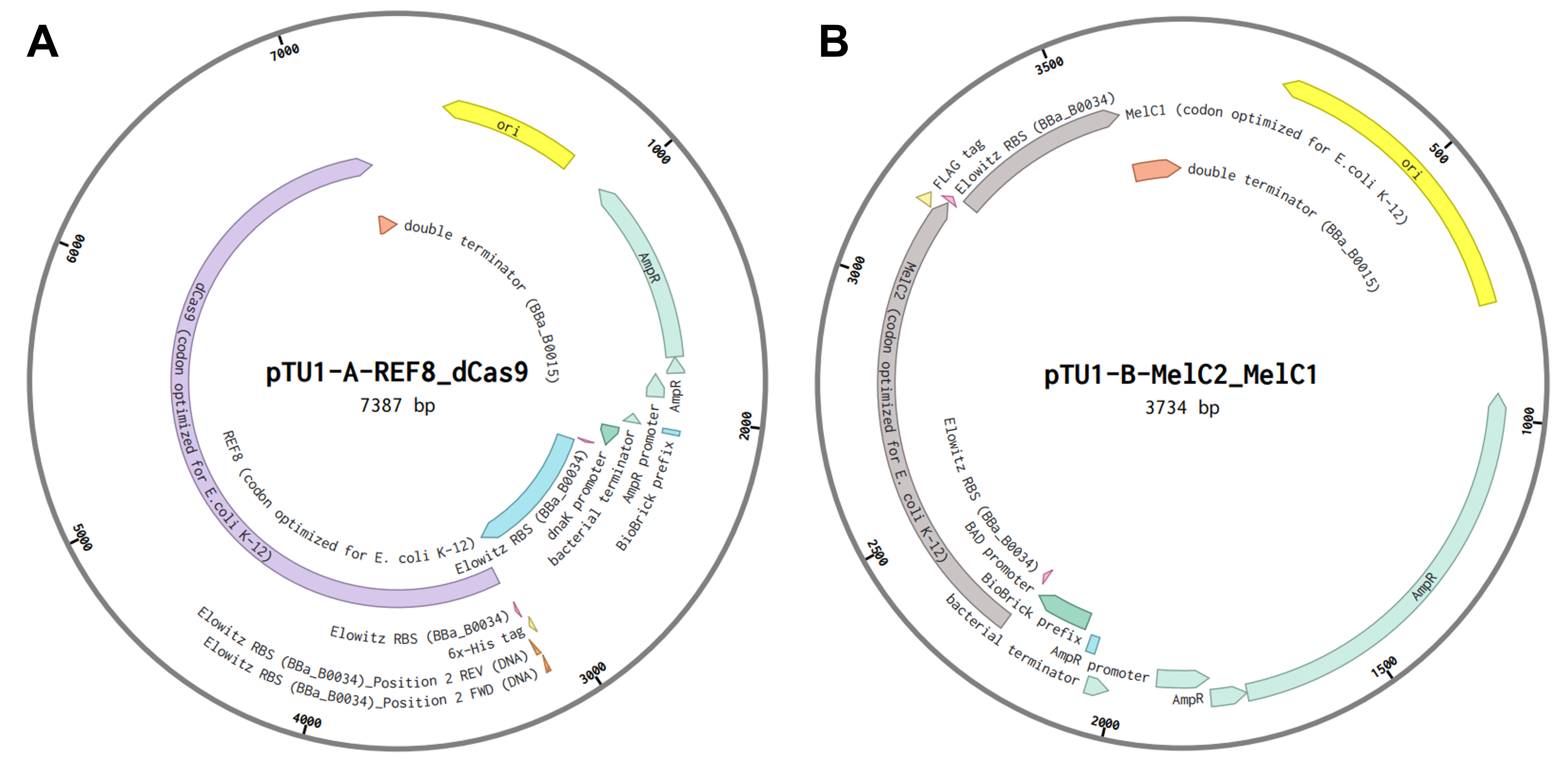
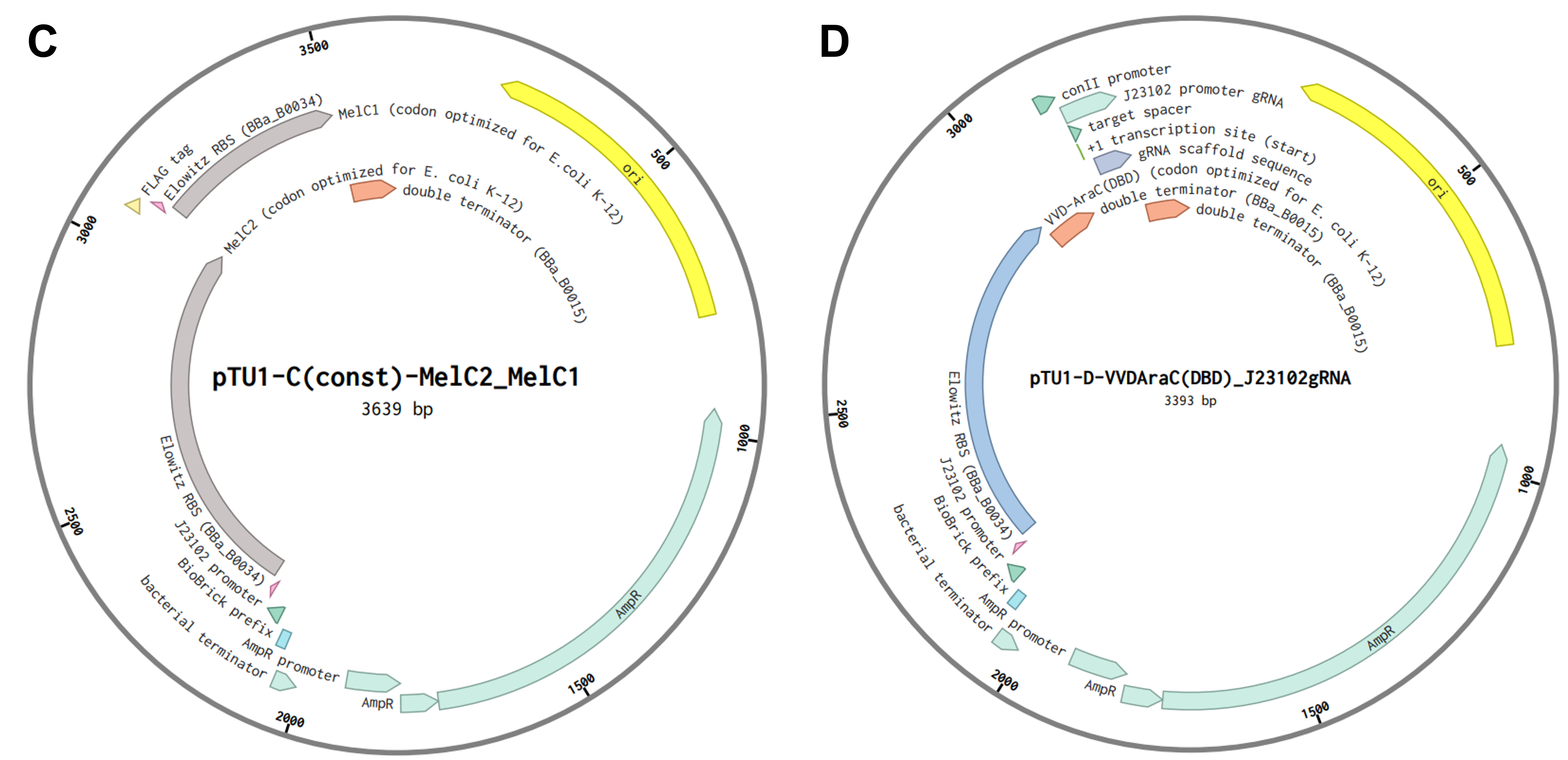
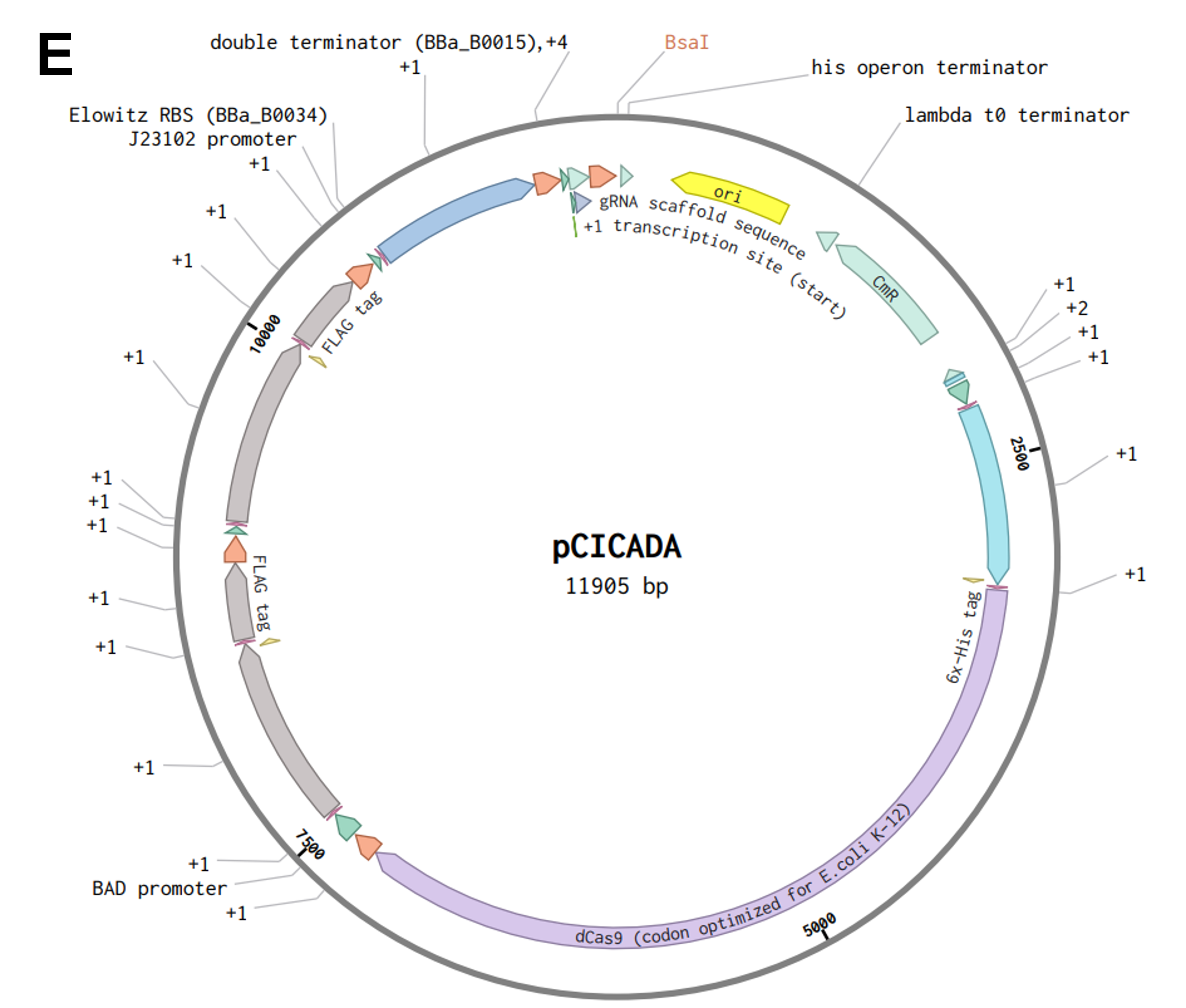 Figure FP.11 Plasmid maps of all level 1 bacterial constructs designed for this project (A - D) and of the final level 2 construct, pCICADA (E), which contains the entire color-changing circuit and will be used for bacterial transformation. Figure created in BioRender.
Figure FP.11 Plasmid maps of all level 1 bacterial constructs designed for this project (A - D) and of the final level 2 construct, pCICADA (E), which contains the entire color-changing circuit and will be used for bacterial transformation. Figure created in BioRender.
Table FP.1 General information about the genetic parts and plasmid backbones utilized for the bacterial constructs.
| Genetic part | Organism of origin | Sequence source |
|---|---|---|
| P dnaK | E. coli (strain K-12, substrain MG1655) | Entry BBa_J100387 in the iGEM Registry of Standard Biological Parts |
| Elowitz RBS | E. coli | Entry BBa_B0034 in the iGEM Registry of Standard Biological Parts |
| REF8 CDS | Sepia officinalis | REF8 CDS submitted on NCBI’s repository 28 |
| dCas9 CDS | Synthetic gene (protein scaffold from Streptococcus pyogenes) | Plasmid pSET-dCas9 in the Addgene repository 46 |
| BBa_B0015 double terminator | Synthetic terminator | Entry BBa_B0015 in the iGEM Registry of Standard Biological Parts |
| P BAD | E. coli and other bacteria | Entry BBa_K206000 in the iGEM Registry of Standard Biological Parts |
| MelC2 CDS | Streptomyces griseus | MelC2 CDS submitted on NCBI’s repository |
| MelC1 CDS | S. griseus | MelC1 CDS submitted on NCBI’s repository |
| J23102 promoter | Synthetic promoter | Entry BBa_J23102 in the iGEM Registry of Standard Biological Parts |
| VVD-AraC(DBD) CDS | Synthetic gene (protein scaffold from E. coli and Neurospora crassa) | Plasmid pBLADE(FP6**)-mCherry in the Addgene repository 38 |
| P conII | Synthetic promoter (promoter scaffold from E. coli) | P conII sequence from Trieu et al., 2026 40 |
| J23102 promoter gRNA | Synthetic gene | Designed with Benchling’s tool for CRISPR-guided interference and partially based on a protocol from the Church Lab |
| pTU1-A-lacZ | Synthetic plasmid | MoClo level 1 plasmid backbone from Moore et al., 2016 45 |
| pTU1-B-lacZ | Synthetic plasmid | MoClo level 1 plasmid backbone from Moore et al., 2016 45 |
| pTU1-C-lacZ | Synthetic plasmid | MoClo level 1 plasmid backbone from Moore et al., 2016 45 |
| pTU1-D-lacZ | Synthetic plasmid | MoClo level 1 plasmid backbone from Moore et al., 2016 45 |
| pTU2-A-RFP | Synthetic plasmid | MoClo level 2 plasmid backbone from Moore et al., 2016 45 |
Step-by-step outline of experiments
After concluding the level 1 and level 2 cloning process as described above and successfully obtaining the pCICADA construct, the next experimental steps are (also compatible the Opentrons/Nebula platform to a large degree):
- Transform competent DH5α E. coli cells with the pCICADA plasmid either through heat shock (exposure to 42°C for 50 seconds - 2 minutes) or electroporation. Plate them in a petri dish, on LB agar with the appropriate selection marker (chloramphenicol, ~25μg/mL) and incubate at 37°C overnight (or for at least 16 hours).
- Inspect the plate after incubation and prepare liquid cultures (again, with chloramphenicol) by inoculating them with bacterial colonies that have emerged. Incubate the liquid cultures at 37°C overnight (or for at least 16 hours).
- Use the bacteria from the liquid cultures to prepare a glycerol stock of E. coli DH5α transformed with pCICADA (for long-term storage of the construct), which will be frozen at -80°C.
- Use the same liquid cultures to extract and purify pCICADA for future transformations.
- Transform competent BL21 E. coli cells with the pCICADA plasmid through heat shock or electroporation, plate them in chloramphenicol-containing LB agar, and incubate them at 37°C overnight.
- Prepare liquid cultures by inoculating them with successfully transformed BL21 colonies. Incubate the bacterial cultures overnight.
- Prior to the induction, normalize cell concentration by isolating roughly the same number of cells for each sample. Prepare biological replicates for each different combination condition (triplicates should be the minimum) for downstream statistical analysis of the experimental results. Controls, probably bacteria that constitutively express reflectin and the L-DOPA biosynthetic enzymes, should be included in triplicates too, for each condition. A pre-culture may also be necessary.
- For the induction, incubate each set of biological replicates and controls under one of four combinations of environmental conditions for approximately two hours: a. at 42°C under constant blue light-enriched illumination, b. at 42°C without illumination, c. at 30 - 37°C under constant blue light-enriched illumination, and d. at 30 - 37°C without illumination. The duration of the induction, the light intensity, and the temperature for the activation of the heat-inducible promoter (42°C) may have to be adjusted for future iterations of the induction experiment depending on the bacteria’s response to the conditions.
- After the induction is complete, normalize cell concentration again to avoid introducing bias to the measurements. Quantify the results of the experiment, namely the levels of reflectin and melanin synthesis, by: a. measuring the absorbance of the induced samples at ~335nm for melanin with a common spectrophotometer and their reflectivity with specialized spectrophotometers optics and optical systems manufacturers (such as Optimax Systems) employ for reflectin; b. visualizing and comparing reflectin and melanin biosynthesis enzyme production across samples through running an SDS-PAGE electrophoresis and subsequently performing a Western blot with anti-His and anti-FLAG antibodies; however, this is a semi-quantifying method that requires isolating and lysing the cells, as well as partially denaturing their proteins, before the protein electrophoresis and the Western blot; c. measuring the actual quantities of synthesized reflectin and L-DOPA melanin/melanin biosynthesis enzymes through mass spectrometry.
- Analyze the obtained results with specialized statistical tools.
Tools and technology
We discussed and practiced various techniques related to synthetic biology throughout the semester. Place a check next to the techniques relevant to your project.
Foundational lab practices
- Pipetting
- Lab safety
- Bioethical considerations (must check this box)
DNA skills and analysis
- DNA gel art
- DNA sequencing
- DNA editing
- DNA construct design
- Restriction enzyme digestion
- Gel electrophoresis
- DNA purification from gel
- Databases (e.g., GenBank, NCBI, Ensembl, and UCSC Genome Browser)
Bioproduction
- Bioproduction
- Chassis selection (e.g., DH5α, Bl21-DE3 for expression)
- Registry of Standard Biological Parts
- Plasmid preparation
- Bacterial culturing
- Quality control/analysis
- Bacterial processing (e.g., centrifugation, lysis, DNA purification)
Lab automation
- Creating code for laboratory automation
- Using liquid handling robots (e.g., Opentrons)
- Designing a Twist order
- Creating a plan to use the Autonomous Lab at Ginkgo Bioworks
Protein design
- Protein design
- Use of Boltz or PepMLM
- Use of Asimov Kernel
- Use of Benchling
- Models and notebooks
- Databases
Cell-free systems
- Cell-free reactions
- Freeze-dried cell-free systems
- NEB Express kits
- miniPCR tools
- Protein purification
Gibson Assembly
- Primer design or selection
- PCR reactions
- Gibson Assembly
- Other cloning methods (e.g., restriction enzyme digestion or Gateway Cloning)
CRISPR
- CRISPR/Cas9
- Designing prime editing gRNA
Section 5: Results and quantitative expectations
As a committed online listener, I did not have access to a lab to conduct experiments and obtain experimental results. However, that did not stop me from speculating what the experimental outcome would be if I exposed bacteria transformed with the pCICADA system to the four different combinations of environmental conditions I listed in my experimental outline. The following imaginary results showcase how the color-changing bacteria would respond to environmental stimuli in an ideal scenario, where regardless of light conditions, high temperatures cause the white coloration to prevail and lower temperatures result in the darker coloration dominating. Figure FP.12 illustrates the results anticipated in the case that the color-shifting circuit works perfectly, along with analytically illustrating the mechanisms and interactions among the circuit’s individual parts that lead to said results. Complementing the qualitative character of Figure FP.12, Figure FP.13 focuses on a more quantitative approach.
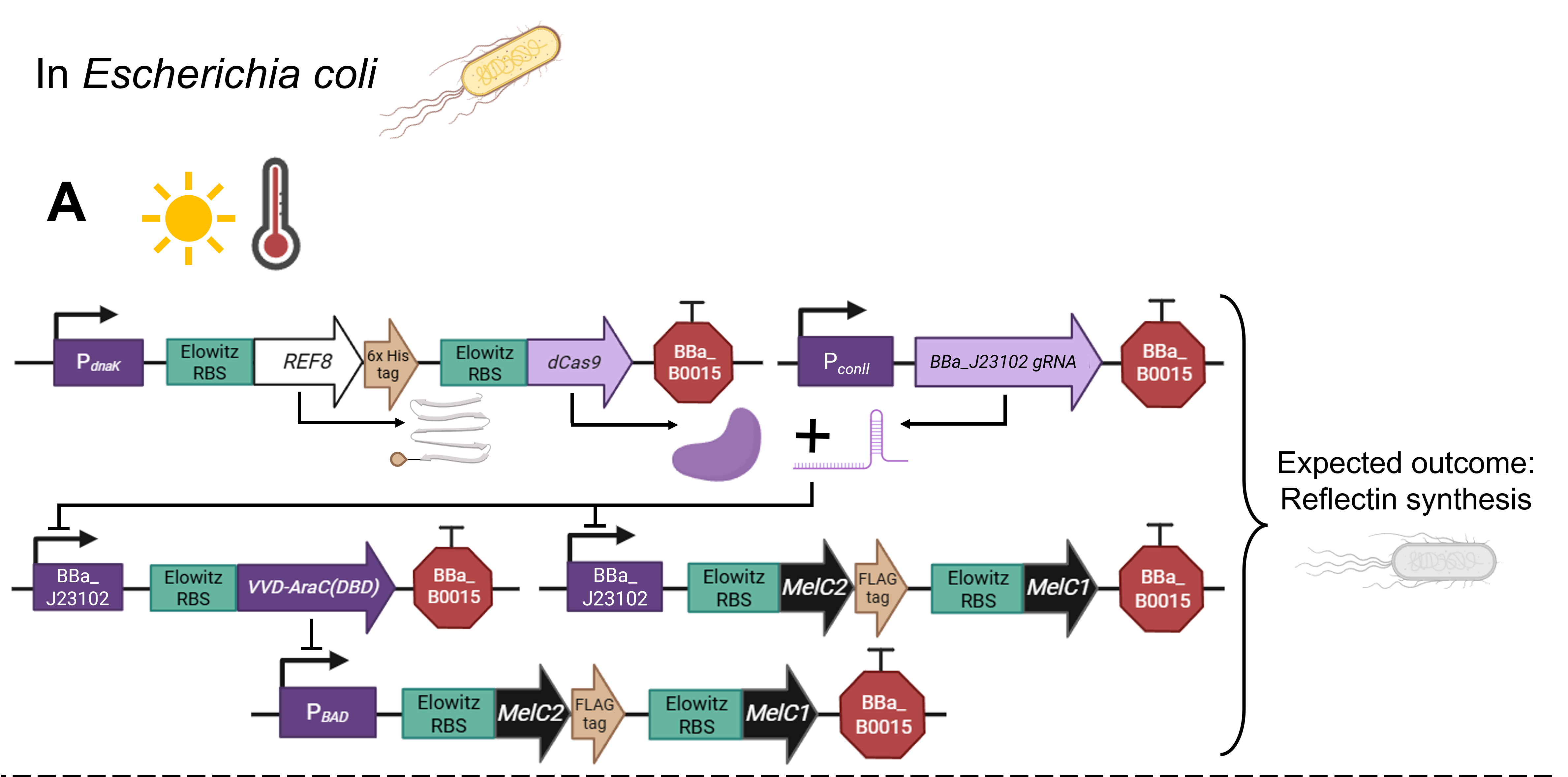
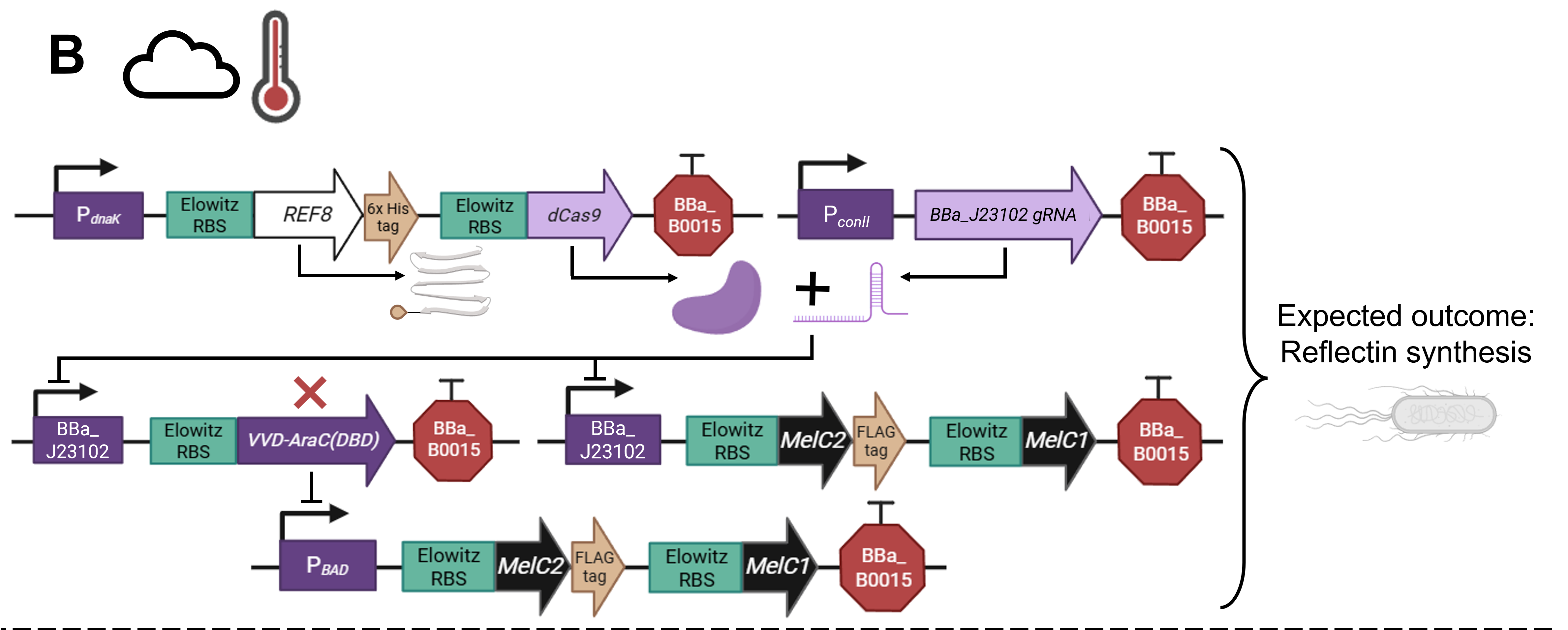
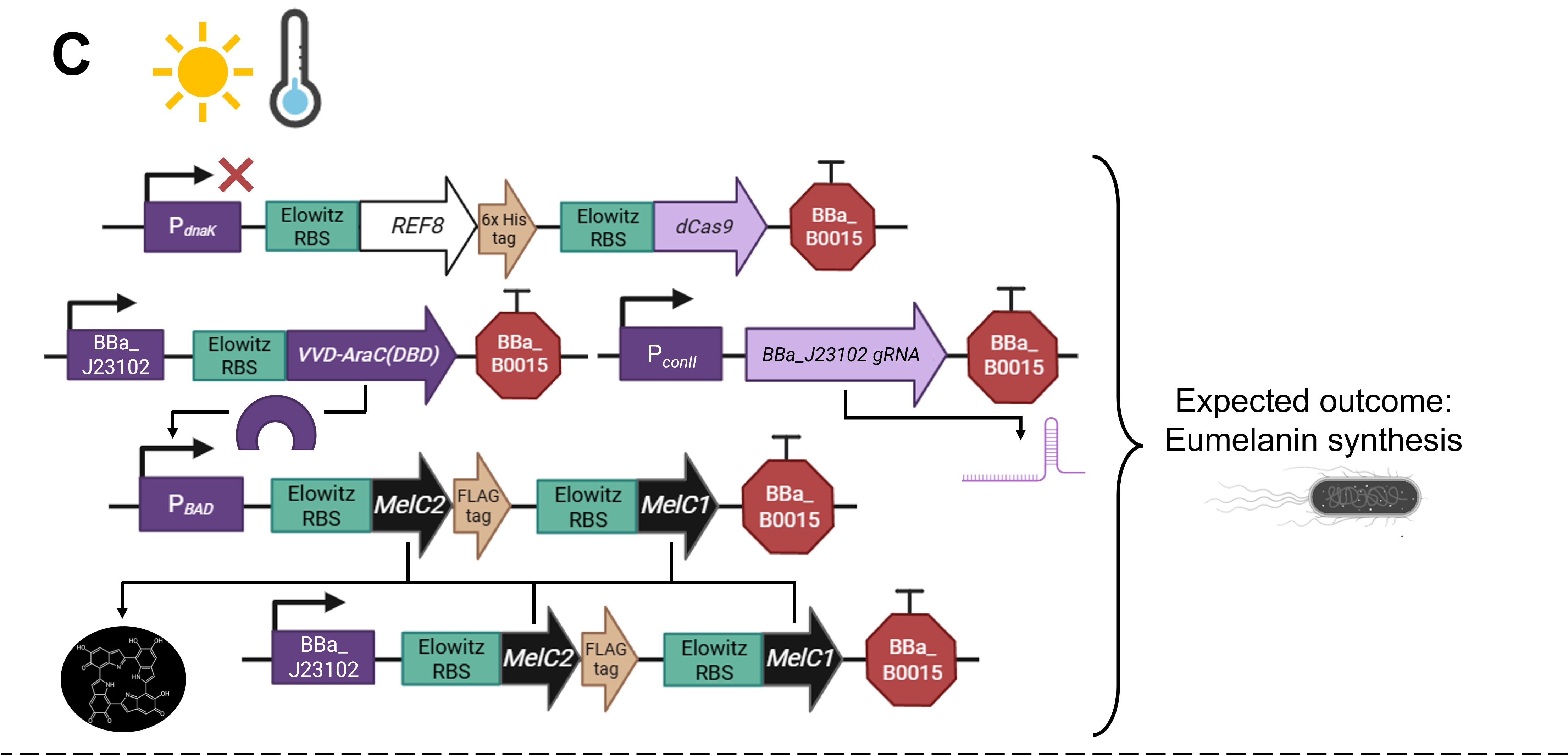
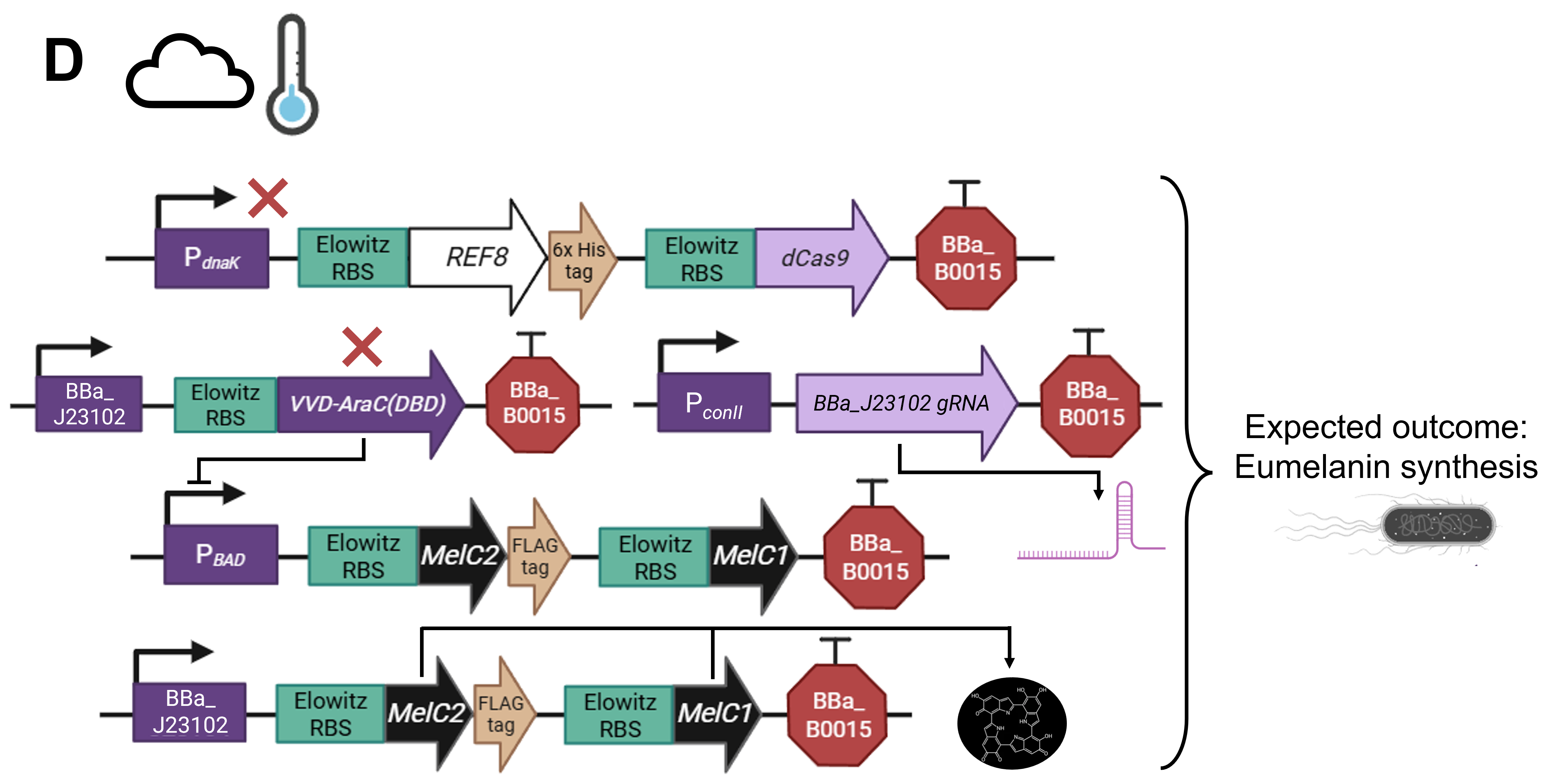 Figure FP.12 Visualization of the anticipated results produced by the activation of the designed pCICADA genetic construct in E. coli under different environmental conditions: (A) under illumination and at a high temperature, (B) in darkness and at a high temperature, (C) under illumination and at a low temperature, and (D) in darkness at a low temperature. Figure created in BioRender.
Figure FP.12 Visualization of the anticipated results produced by the activation of the designed pCICADA genetic construct in E. coli under different environmental conditions: (A) under illumination and at a high temperature, (B) in darkness and at a high temperature, (C) under illumination and at a low temperature, and (D) in darkness at a low temperature. Figure created in BioRender.
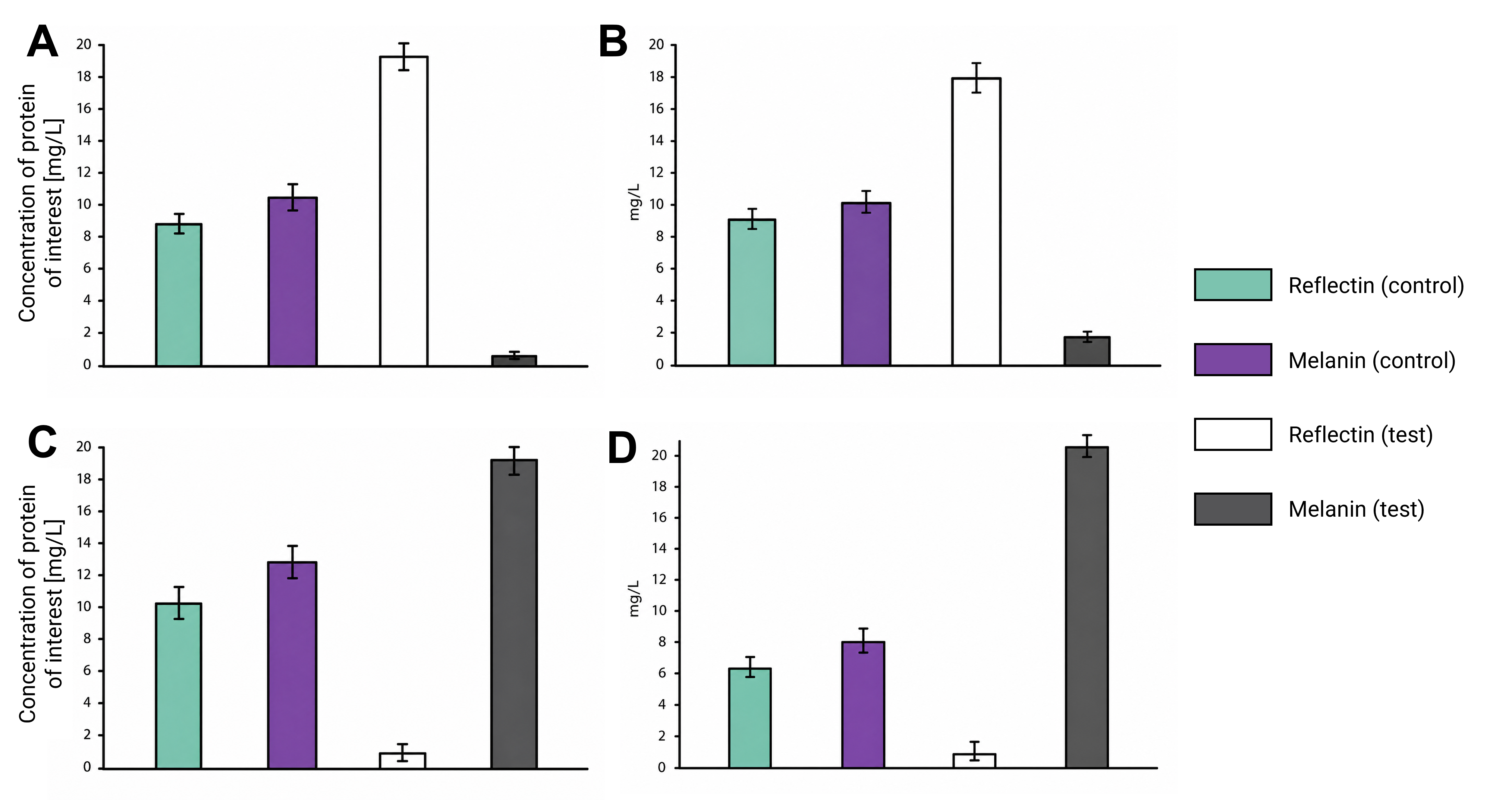 Figure FP.13 Graphs showing spectrophotometric data-based predicted measurements generated by the activation of the pCICADA construct in E. coli under different environmental conditions: (A) under illumination and at a high temperature, (B) in darkness and at a high temperature, (C) under illumination and at a low temperature, and (D) in darkness at a low temperature.
Figure FP.13 Graphs showing spectrophotometric data-based predicted measurements generated by the activation of the pCICADA construct in E. coli under different environmental conditions: (A) under illumination and at a high temperature, (B) in darkness and at a high temperature, (C) under illumination and at a low temperature, and (D) in darkness at a low temperature.
Section 6: Discussion and future work
- See also the aims at the beginning
- Pilot implementation of the project after appropriate biosecurity considerations
- Neuromorphic circuit
- Alternative pathways of melanin production in Aspergillus? More copies of the PKS1 gene?
- Promoter engineering to render the heat- and light-inducible promoters capable of being activated at 25 to 30 degrees and by more wavelenght frequencies in the visible light spectrum
- Optimized fungal construct
- Insulator elements
- Ethical consideration: non-malevolent use of the biomaterial, sustainability and respect to ecosystem, biosecurity mechanisms
- Application method (figure below)
Section 7: Additional information
Shokoohi K. The ancient Persian way to keep cool. Published February 24, 2022. https://www.bbc.com/future/article/20210810-the-ancient-persian-way-to-keep-cool ↩︎
Bock P. Singapore’s Oasia Hotel Downtown has its own ecosystem. WIRED. https://www.wired.com/story/oasia-singapore-ecosystem-hotel/. Published March 12, 2017. ↩︎
DECA architecture. https://deca.gr/ ↩︎
DECA architecture’s new office in an old industrial building | Transsolar | KlimaEngineering. Transsolar | KlimaEngineering. https://transsolar.com/projects/athen-deca-office ↩︎ ↩︎
Hanlon R. Cephalopod dynamic camouflage. Curr Biol. 2007;17(11):R400-4. doi:10.1016/j.cub.2007.03.034 ↩︎ ↩︎
Amodio P, Boeckle M, Schnell AK, Ostojíc L, Fiorito G, Clayton NS. Grow smart and die young: why did cephalopods evolve intelligence? Trends Ecol Evol. 2019;34(1):45-56. doi:10.1016/j.tree.2018.10.010 ↩︎
Gonzalez-Bellido PT, Scaros AT, Hanlon RT, Wardill TJ. Neural Control of Dynamic 3-Dimensional Skin Papillae for Cuttlefish Camouflage. iScience. 2018;1:24-34. doi:10.1016/j.isci.2018.01.001 ↩︎
Mäthger LM, Denton EJ, Marshall NJ, Hanlon RT. Mechanisms and behavioural functions of structural coloration in cephalopods. J R Soc Interface. 2009;6 Suppl 2(Suppl 2):S149-63. doi:10.1098/rsif.2008.0366.focus ↩︎ ↩︎ ↩︎
Williams TL, Senft SL, Yeo J, et al. Dynamic pigmentary and structural coloration within cephalopod chromatophore organs. Nat Commun. 2019;10(1):1004. doi:10.1038/s41467-019-08891-x ↩︎ ↩︎
Lewis LLM, Dörschmann P, Seeba C, et al. Properties of Cephalopod Skin Ommochromes to Inhibit Free Radicals, and the Maillard Reaction and Retino-Protective Mechanisms in Cellular Models Concerning Oxidative Stress, Angiogenesis, and Inflammation. Antioxidants (Basel). 2022;11(8). doi:10.3390/antiox11081574 ↩︎
Dessi-Olive J. Strategies for Growing Large-Scale Mycelium Structures. Biomimetics (Basel). 2022;7(3). doi:10.3390/biomimetics7030129 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Li K, Wei Z, Jia J, et al. Engineered living materials grown from programmable Aspergillus niger mycelial pellets. Materials Today Bio. 2023;19:100545. doi:10.1016/j.mtbio.2023.100545 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Globa A, Soh E, Le Ferrand H. Living Textures and Mycelium Skin Co-Creation: Designing Colour, Pattern, and Performance for Bio-Aesthetic Expression in Mycelium-Bound Composites. Biomimetics (Basel). 2025;10(9). doi:10.3390/biomimetics10090573 ↩︎
Guarino B. Researchers invent camouflaged membrane that hides like an octopus. The Washington Post. https://www.washingtonpost.com/news/speaking-of-science/wp/2017/10/12/researchers-invent-camouflaged-membrane-that-hides-like-an-octopus/. Published October 27, 2021. ↩︎
Morillas L, Roales J, Cruz C, Munzi S. Lichen as multipartner symbiotic relationships. Encyclopedia. 2022;2(3):1421-1431. doi:10.3390/encyclopedia2030096 ↩︎ ↩︎ ↩︎ ↩︎
Rokas A. Aspergillus. Current Biology. 2013;23(5):R187-R188. doi:10.1016/j.cub.2013.01.021 ↩︎ ↩︎
Oakley BR. Aspergillus nidulans. In: Elsevier eBooks. ; 2016. doi:10.1016/b978-0-12-809633-8.06093-3 ↩︎
Ghazawi KF, Fatani SA, Mohamed SGA, Mohamed GA, Ibrahim SRM. Aspergillus nidulans—Natural Metabolites Powerhouse: Structures, Biosynthesis, Bioactivities, and Biotechnological Potential. Fermentation. 2023;9(4):325. doi:10.3390/fermentation9040325 ↩︎
Krespach MKC, García-Altares M, Flak M, et al. Lichen-like association of Chlamydomonas reinhardtii and Aspergillus nidulans protects algal cells from bacteria. ISME J. 2020;14(11):2794-2805. doi:10.1038/s41396-020-0731-2 ↩︎
Jiang L, Li T, Jenkins J, et al. Evidence for a mutualistic relationship between the cyanobacteria Nostoc and fungi Aspergilli in different environments. Appl Microbiol Biotechnol. 2020;104(14):6413-6426. doi:10.1007/s00253-020-10663-3 ↩︎
Chhoker K, Hausner G, Harris SD. Regulation of melanin production in fungi. Front Fungal Biol. 2025;6:1621764. doi:10.3389/ffunb.2025.1621764 ↩︎
Gonçalves RCR, Lisboa HCF, Pombeiro-Sponchiado SR. Characterization of melanin pigment produced by Aspergillus nidulans. World J Microbiol Biotechnol. 2012;28(4):1467-1474. doi:10.1007/s11274-011-0948-3 ↩︎
Medeiros WB, Medina KJD, Sponchiado SRP. Improved natural melanin production by Aspergillus nidulans after optimization of factors involved in the pigment biosynthesis pathway. Microb Cell Fact. 2022;21(1):278. doi:10.1186/s12934-022-02002-0 ↩︎
Jones RD, Qian Y, Siciliano V, et al. An endoribonuclease-based feedforward controller for decoupling resource-limited genetic modules in mammalian cells. Nat Commun. 2020;11(1):5690. doi:10.1038/s41467-020-19126-9 ↩︎
Panchapakesan SSS, Ferguson ML, Hayden EJ, Chen X, Hoskins AA, Unrau PJ. Ribonucleoprotein purification and characterization using RNA Mango. RNA. 2017;23(10):1592-1599. doi:10.1261/rna.062166.117 ↩︎
Kong KJ, Lu X, Dolgosheina E, et al. Fluorogenic aptamers for imaging and manipulation of cellular RNAs. Biophysical Journal. 2022;121(3):318a-319a. doi:10.1016/j.bpj.2021.11.1168 ↩︎
Matsushita M, Tada S, Suzuki S, Kusumoto KI, Kashiwagi Y. Deletion analysis of the promoter of Aspergillus oryzae gene encoding heat shock protein 30. Journal of Bioscience and Bioengineering. 2009;107(4):345-351. doi:10.1016/j.jbiosc.2008.11.024 ↩︎
Bassaglia Y, Bekel T, Da Silva C, et al. ESTs library from embryonic stages reveals tubulin and reflectin diversity in Sepia officinalis (Mollusca — Cephalopoda). Gene. 2012;498(2):203-211. doi:10.1016/j.gene.2012.01.100 ↩︎ ↩︎
Unkles SE, Valiante V, Mattern DJ, Brakhage AA. Synthetic biology tools for bioprospecting of natural products in eukaryotes. Chem Biol. 2014;21(4):502-508. doi:10.1016/j.chembiol.2014.02.010 ↩︎
Schuetze T, Meyer V. Polycistronic gene expression in Aspergillus niger. Microb Cell Fact. 2017;16(1):162. doi:10.1186/s12934-017-0780-z ↩︎
Hernández G, Osnaya VG, Pérez-Martínez X. Conservation and variability of the AUG initiation codon context in eukaryotes. Trends Biochem Sci. 2019;44(12):1009-1021. doi:10.1016/j.tibs.2019.07.001 ↩︎
Hedtke M, Rauscher S, Röhrig J, Rodríguez-Romero J, Yu Z, Fischer R. Light-dependent gene activation in Aspergillus nidulans is strictly dependent on phytochrome and involves the interplay of phytochrome and white collar-regulated histone H3 acetylation. Mol Microbiol. 2015;97(4):733-745. doi:10.1111/mmi.13062 ↩︎
Yu Z, Streng C, Seibeld RF, et al. Genome-wide analyses of light-regulated genes in Aspergillus nidulans reveal a complex interplay between different photoreceptors and novel photoreceptor functions. PLoS Genet. 2021;17(10):e1009845. doi:10.1371/journal.pgen.1009845 ↩︎
Chavez C, Cruz-Becerra G, Fei J, Kassavetis GA, Kadonaga JT. The tardigrade damage suppressor protein binds to nucleosomes and protects DNA from hydroxyl radicals. eLife. 2019;8. doi:10.7554/eLife.47682 ↩︎
Son Y-E, Park H-S. Genetic Manipulation and Transformation Methods for Aspergillus spp. Mycobiology. 2021;49(2):95-104. doi:10.1080/12298093.2020.1838115 ↩︎
Elowitz MB, Leibler S. A synthetic oscillatory network of transcriptional regulators. Nature. 2000;403(6767):335-338. doi:10.1038/35002125 ↩︎
Seaton BL, Vickery LE. A gene encoding a DnaK/hsp70 homolog in Escherichia coli. Proc Natl Acad Sci USA. 1994;91(6):2066-2070. doi:10.1073/pnas.91.6.2066 ↩︎
- Romano E, Baumschlager A, Akmeriç EB, et al. Engineering AraC to make it responsive to light instead of arabinose. Nat Chem Biol. 2021;17(7):817-827. doi:10.1038/s41589-021-00787-6
Pavan ME, López NI, Pettinari MJ. Melanin biosynthesis in bacteria, regulation and production perspectives. Appl Microbiol Biotechnol. 2020;104(4):1357-1370. doi:10.1007/s00253-019-10245-y ↩︎
Trieu KP, Bishé B, Taton A, Tieu BP, Golden JW. A set of constitutive promoters with graded strengths for gene expression in diverse cyanobacterial strains. BioRxiv. March 26, 2026. doi:10.64898/2026.03.25.714268 ↩︎ ↩︎
Elledge SJ, Davis RW. Position and density effects on repression by stationary and mobile DNA-binding proteins. Genes Dev. 1989;3(2):185-197. doi:10.1101/gad.3.2.185 ↩︎
Didovyk A, Borek B, Tsimring L, Hasty J. Transcriptional regulation with CRISPR-Cas9: principles, advances, and applications. Curr Opin Biotechnol. 2016;40:177-184. doi:10.1016/j.copbio.2016.06.003 ↩︎
Anderson DA, Voigt CA. Competitive dCas9 binding as a mechanism for transcriptional control. Mol Syst Biol. 2021;17(11):e10512. doi:10.15252/msb.202110512 ↩︎
Hawkins JS, Wong S, Peters JM, Almeida R, Qi LS. Targeted transcriptional repression in bacteria using CRISPR interference (crispri). Methods Mol Biol. 2015;1311:349-362. doi:10.1007/978-1-4939-2687-9_23 ↩︎
Moore SJ, Lai HE, Kelwick RJ, et al. Ecoflex: A multifunctional MoClo kit for E. coli synthetic biology. ACS Synth Biol. 2016;5(10):1059-1069. doi:10.1021/acssynbio.6b00031 ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎ ↩︎
Zhao Y, Li L, Zheng G, et al. CRISPR/DCAS9‐Mediated Multiplex Gene Repression in streptomyces. Biotechnology Journal. 2018;13(9):e1800121. doi:10.1002/biot.201800121 ↩︎