Individual Final Project
HTGAA 2026: Individual Final Project Documentation
SECTION 1: ABSTRACT
This project develops a computational workflow for the rational design of binders targeting ferritin heavy chain (FRIH), an iron storage protein for which no minimally invasive biosensor currently exists. Using RNAfold secondary structure analysis, 100 candidate ssDNA sequences (40 nt) were screened and filtered by minimum free energy criteria. A proximity ligation assay (PLA) framework was proposed to de-risk molecular recognition before device integration, combining a Nb72 nanobody capture probe with a FRIH-directed DNA aptamer reporter in an electrochemical detection architecture. The workflow is broadly adaptable and can be optimized for other clinically important biomarkers, including inflammatory proteins (CRP, IL-6) and cardiac markers (NT-proBNP), supporting a scalable route from computational design to clinical biosensor deployment.
SECTION 2: PROJECT AIMS
Computational design and prioritization.
Identify known binders (nanobodies, DNA aptamers) of the human ferritin heavy chain by literature and database searches. If no binders existed, computationally design and validate de novo binders. By using secondary structure analysis tools such as RNAfold, and for 3D structure prediction by RNAComposer. Finally, validate the binders by molecular docking at the protein interface.
Although the approach here is computational, an experimental approach is the gold standard, such as performing SELEX (Systematic Evolution of Ligands by Experimental Enrichment) for the identification of DNA aptamers.
Design a DNA construct for cell-free protein synthesis to express and purify the human ferritin heavy chain. Validate the DNA construct in silico for the expected outcome by restriction enzyme digestion. Place an order for the DNA construct to be synthesized by Twist Bio.
Below are the potential resources for rational binder design:
De novo DNA aptamer design using bioinformatics.org (random DNA generator), NUPACK, RNAfold, RNAComposer, Mfold, HADDOCK 2.4.
De novo nanobody design using RFantibody, BindCraft, AlphaFold2, AlphaFold3, ESM2, and ProteinMPNN.
Protein docking using AlphaFold3, HADDOCK 2.4.
3D structure visualization tool using UCSF ChimeraX.
Aim 2: Development Aim
Validation with lab assays and microneedle array fabrication.
DNA aptamers identified by computational tools can be validated in various laboratory assays, such as in vitro assays, SELEX, and in vivo assays, such as the yeast-surface-display assay.
A cell-free protein synthesis expression system can be validated by protein expression and His tag affinity purification. Ferritin is a 24-mer protein. The oligomeric state of the purified protein can be validated by charge state mass spectroscopy (CSMS) analysis. These validation experiments can potentially be run in Gingko cloud labs (Nebula), and purified protein can be tested for oligomeric state at Waters (Xevo CDMS).
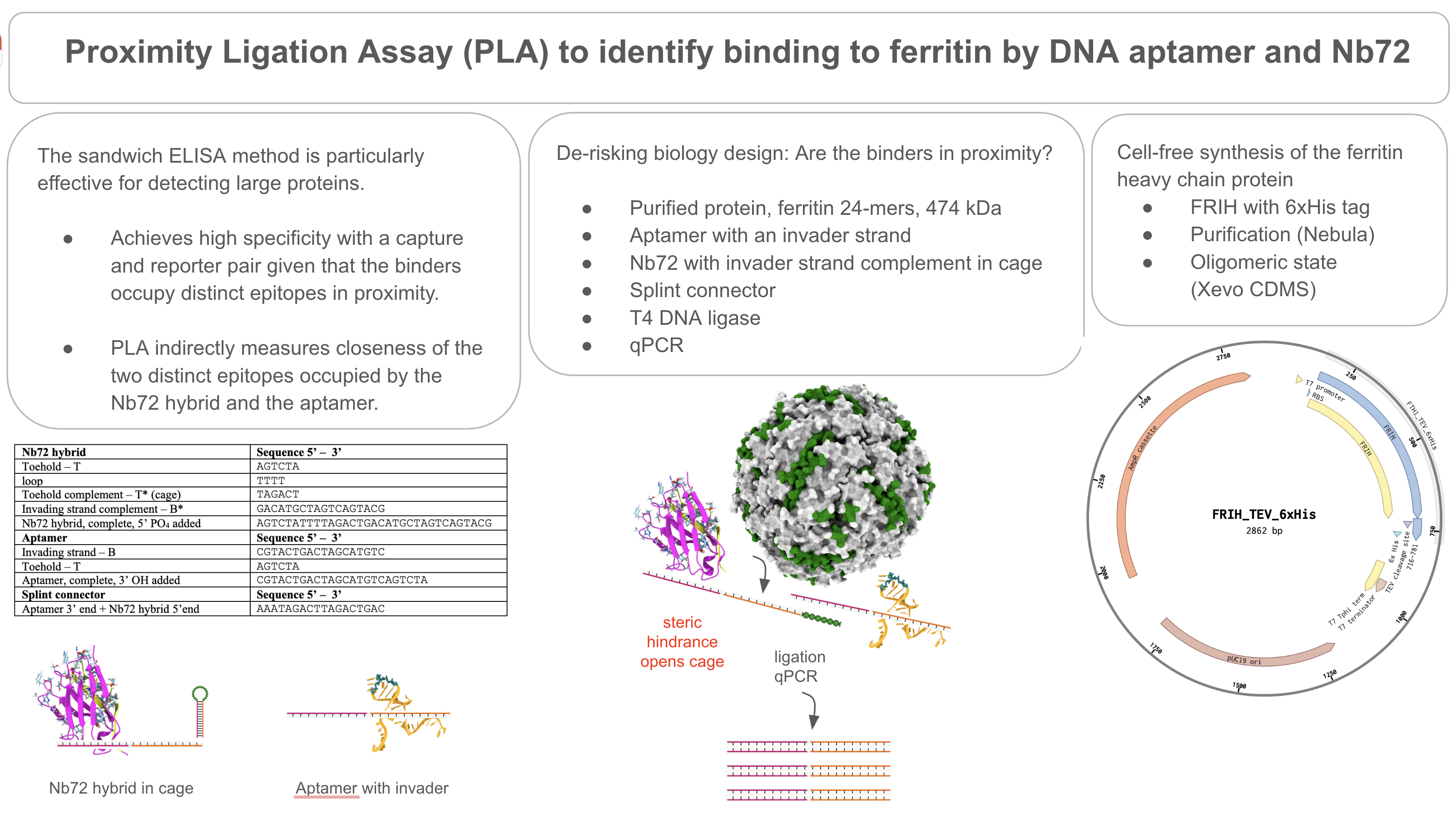
Biology design should be de-risked before microneedle array fabrication. Epitope binding, proximity of binding sites, and the design of extended arm sequences (a caged toehold and an invader strand) can be validated in a proximity ligation assay (PLA) with a qPCR readout.
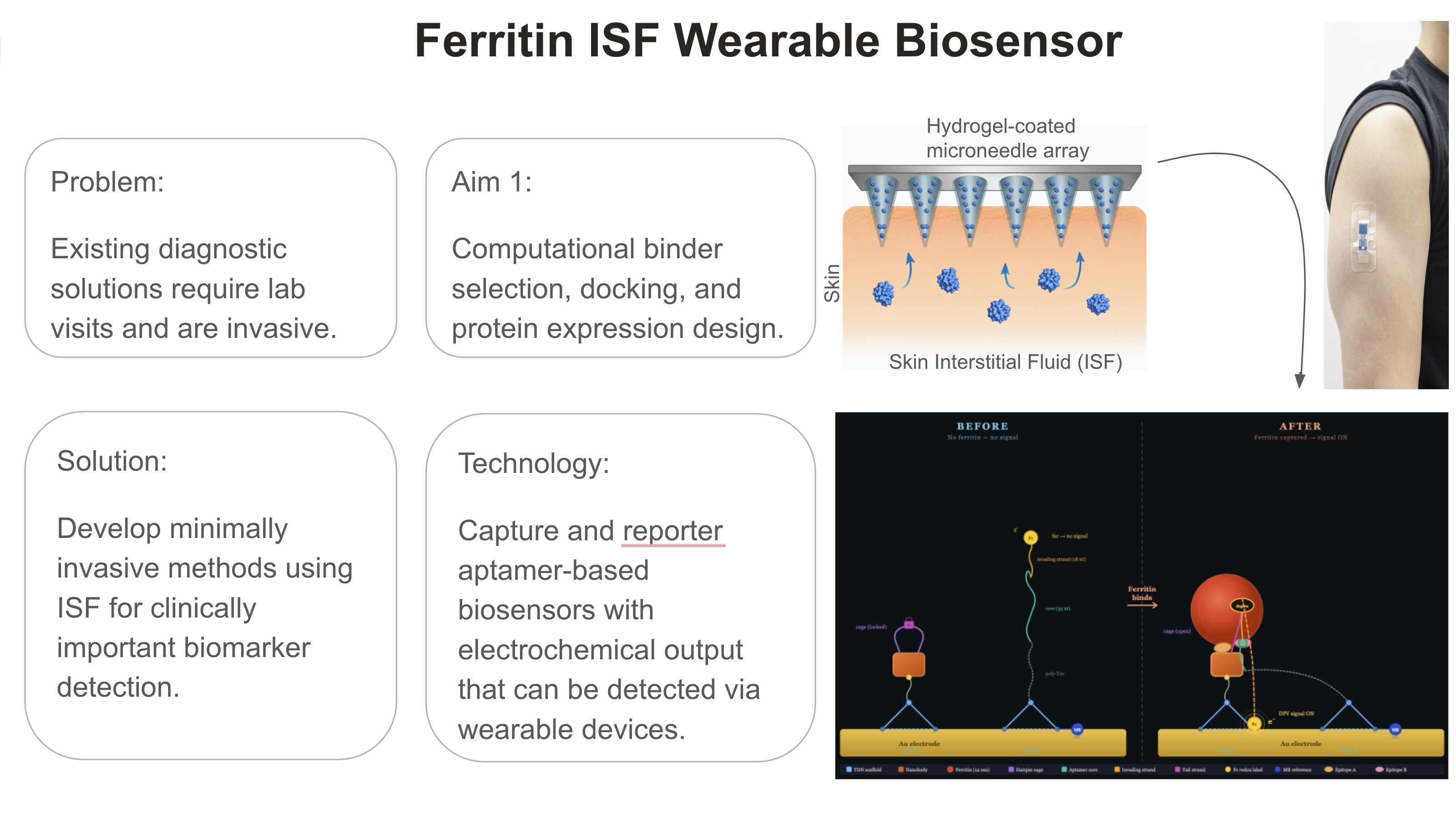
Microneedle array fabrication. All biology components, the nanobody binder and the DNA aptamer, will be assembled on a gold electrode that generates an electrochemical readout. The system can be validated by limit of detection (LoD) studies with artificial ISF spiked with purified ferritin protein.
Aim 3: Visionary Aim
Clinical trials, blood correlations, and validations.
Minimally invasive diagnostics based on ISF are at a transformative inflection point. While continuous glucose monitors have established ISF as a viable diagnostic matrix, the field is now positioned to expand toward continuous, multiplexed monitoring of a broad panel of clinically important biomarkers. This project develops a biosensor platform for ISF ferritin detection, establishing design principles that are generalizable to other targets. Computational aptamer design tools and proximity ligation-based screening and validation provide a scalable and cost-effective pathway to accelerate biosensor development toward clinically deployable wearable devices.
The platform architecture is inherently modular; the recognition element can be exchanged to redirect the biosensor toward any protein target. This positions the platform as a versatile foundation for detecting clinically important ISF-accessible biomarkers, including inflammatory proteins such as CRP and IL-6, and cardiac markers such as NT-proBNP.
The clinical and population-level impact of this platform is substantial. At the individual level, continuous ISF biomarker monitoring enables early disease detection and real-time tracking of therapeutic response, reducing patient burden and enabling personalized care without repeated hospital visits. At the population level, the lab-on-chip format is operational without specialized infrastructure or trained personnel. This makes large-scale diagnostic deployment feasible in resource-limited, remote, and underserved settings. This democratization of diagnostic access shifts the healthcare paradigm from reactive, symptom-driven encounters toward proactive, data-driven health monitoring across diverse populations.
SECTION 3: BACKGROUND
Background and Literature Context.
Highly desirable personalized healthcare innovations for unmet clinical needs have driven accelerated developments in the minimally invasive diagnostics field in recent years. There is a real value in reducing patient burden and enabling continuous or real-time biomarker monitoring when it comes to disease prevention and improving the quality of patient lives. Adding to this paradigm, reducing cost to enable broad-population-level accessibility. Traditional diagnostic care generally relies on a laboratory visit, blood withdrawal, trained personnel, and a high cost for housing instruments and maintenance, and trained professionals to analyze the results. On the other hand, wearable monitoring devices capable of detecting and translating specific biological signals at the point-of-care would overcome these limitations. Continuous glucose monitors in diabetes management are widely recognized as a success in the minimally invasive diagnostics field, and a major motivator for extending to include a broader panel of clinical biomarkers accessible from dermal interstitial fluid (ISF) (Rodbard 2016, Klonoff et al. 2017, Liang et al. 2024). Although advances have been made, as demonstrated with the development of caffeine, levodopa, and beta-lactam antibiotic detection systems, there are challenges around detection limits and validation (blood vs ISF) (Sprunger et al. 2025).
Interstitial fluid is the fluid surrounding cells in peripheral tissues. It is the medium for nutrient delivery, waste removal, and molecular signaling. It constitutes approximately 75% of extracellular fluid, making it one of the most diagnostically accessible fluids given its proximity to skin surface. Studies performed by several groups suggest that protein concentrations in ISF fall between 50-60% plasma levels. On average, the ISF/serum total protein ratio is 57% (Sprunger et al. 2025). Metabolite profiling reported by another study indicates that 68 % of detected metabolite features were shared between plasma and skin dialysate, and significant positive correlations were found across 39 metabolites between the two matrices. (Oharazawa et al. 2024). These studies also point out that some analytes are locally produced in skin tissue and therefore uniquely present in ISF, a key testing advantage of ISF. Sprunger et al screened over 50 analytes plus the Olink panel (48 cytokines) in healthy individuals and found that of the Olink panel specifically, 36 of 45 were over the limit of quantification in both ISF and serum. Notably, the authors demonstrated the detectability of NT-proBNP, a cardiac failure biomarker at comparable concentrations in serum and ISF (Sprunger et al. 2025), indicating ISF can serve as a reliable proxy for monitoring cardiac health.
Data support a rich composition of ISF and the detectability of biomarkers. However, ISF diagnostics face important challenges. As reported by Sprunger et al., there is a relationship between the molecular weight and serum-to-ISF dilution factor (R2=0.87), which is consistent with the hypothesis that passive molecular transport and partitioning between blood and ISF are size-dependent. As projected, size-dependent concentration in ISF has direct implications for the biosensor design that requires enabling the detection platforms to achieve higher sensitivity. This challenge is particularly notable for high-molecular-weight targets such as ferritin, which would be expected to show substantial ISF dilution.
Ferritin is the primary intracellular iron storage protein, is the most reliable serum biomarker for evaluating the body’s iron reserves, and is routinely used to diagnose iron deficiency anemia, liver disease, and iron overload conditions (Vonalioglu et al. 2025). Beyond iron status, ferritin has other clinical relevance. Markedly elevated levels of ferritin correlate with SARS-CoV-2 infection and cytokine storm severity. Additionally, ferritin dysregulation is implicated in neurodegenerative conditions, including Parkinson’s and Alzheimer’s disease.
Traditional ferritin detection relies on ELISA and radioimmunoassays (RIA), detecting a normal concentration range in serum, 15-300 ng/mL, which, while effective, are costly, require specialized materials and trained personnel, and are impractical for point-of-care deployment. This has motivated the development of biosensor-based alternatives, such as graphene-based Field-Effect-Transistor (FET) biosensors, microfluidic electrochemical immunosensors, Surface-Plasmon Resonance (SPR)-based sandwich assays, and paper-based electrochemical devices (Vonalioglu et al. 2025). These alternative platforms achieve varied levels of detection limits as low as 0.19 ng /mL (paper-based) (Boonkaew et al. 2020) and mostly remain at early proof-of-concept stages.
As expected, the development of an ISF ferritin biosensor would face challenges due to its large molecular weight. A hollow cage protein, consisting of 24 subunits, has an approximate molecular mass of 474 kDa (Lawson et al. 1991). Given the molecular size, ferritin is expected to be diluted in ISF. This implies that any ferritin biosensor targeting ISF must achieve substantially lower detection limits than those optimized for serum and must employ recognition elements with high binding affinity to capture the target at low concentrations.
Sandwich immunoassays effectively detect large proteins and are the most clinically validated analytical assays. The format is robust, owing to its dual-epitope specificity requirement, which reduces non-specific signal, and to the signal amplification provided by the secondary detection step. The implication of sandwich-based assays in ferritin detection has been documented (Vonalioglu et al. 2025). However, a key limitation in the sandwich format for ISF deployment is to accommodate the small sample volumes achievable by microneedle collection. These practical constraints motivate the exploration of other biosensor design architectures (Han et al. 2010).
Aptamer-based biosensors are promising alternatives as opposed to antibodies, particularly in terms of thermal stability, sensor regeneration, to be chemically modified with redox labels for electrochemical readout and compatibility with nucleic acid signal amplification architectures. Cost-effective and scalable, aptamers are the single-stranded DNA or RNA molecules selected to bind specific targets with high affinity and specificity (Han et al. 2010).
Several aptamer-based biosensor designs are relevant for detecting proteins from low-concentration samples, such as ISF. A target-induced structure switching (TISS) and a closely related toehold-mediated strand displacement (TMSD) reactions are mechanisms by which target-binding induces conformation change in the aptamer, resulting in the repositioning of an attached redox label relative to an electrode surface and generating an electrochemical signal. A biosensor for thrombin detection described by Xiao et al. is based on TISS (Xiao et al.2005). Wang et al. describe a related approach for ATP detection using a hidden toehold design, in which ATP binding liberates a hidden toehold domain, triggering TMSD on an electrode surface to produce an electrochemical readout (Wang et al. 2015).
Another relevant design is the proximity ligation assay (PLA) described by Fredriksson et al. (Fredriksson et al 2002). In PLA, two recognition probes, aptamers, nanobodies, or antibodies, each conjugated to an oligonucleotide arm, bind distinct epitopes on the same target protein simultaneously. Upon binding to the target, the two arms are brought into proximity, enabling hybridization and subsequent ligation to generate a unique amplifiable DNA sequence. The ligation product is detected via nucleic acid amplification. This design is particularly powerful for large proteins such as ferritin, because it presents numerous accessible epitopes across its surface. and converts a protein-binding event into a nucleic acid amplification event, enabling detection at very low concentrations.
Together, these molecular design strategies, particularly when coupled to electrochemical transduction, offer sensitive and potentially regenerable biosensors for large protein targets in ISF. This project aims to bridge the gap between ISF biomarker accessibility and sensitive electrochemical detection of ferritin using one or more of the molecular strategies reviewed.
Summary of two peer-reviewed research citations relevant to this project.
Paper 1. Sprunger, Y., Longo, J., Saeidi, A., Ionescu, A.M. 2025. Bridging Blood and Skin: Biomarker Profiling in Dermal Interstitial Fluid (dISF) for Minimally Invasive Diagnostics. Biosensors.
A key finding in the Sprunger et al. paper is that large proteins are detectable in ISF but at reduced concentrations relative to blood. They quantified the analytes’ serum-to-ISF partitioning and their molecular weight, demonstrating a strong linear relationship with R² = 0.87. Their result demonstrated that ISF closely resembles serum but at reduced concentrations (57% of total protein). They observed a strong correlation between ISF and serum for CRP and SAA in individuals with pathological levels. Notably, they demonstrated the detectability of NT-proBNP (a cardiac failure biomarker) at comparable levels in both ISF and serum, indicating ISF could serve as a reliable proxy for monitoring cardiac health.
Sprunger et al. compared ISF and serum from volunteers in two separate trials. The first trial was with 12 volunteers and quantified total protein, HSA, and IgG. The second was with 20 volunteers, quantifying 50 inflammation-related protein biomarkers. They used a collection method, a microneedle device, on the biceps-triceps region, recovering an average of 3.4 uL/h without lasting skin damage.
Paper 2. Wang, L., Fang, L., Liu, S. 2015. Responsive Hairpin DNA Aptamer Switch to Program the Strand Displacement Reaction for the Enhanced Electrochemical Assay of ATP. Analyst.
In this paper, the authors designed a hairpin DNA aptamer switch for enzyme-free, electrochemical detection of ATP. A structural change occurs upon ATP binding to the aptamer and subsequently executing the branch migration reaction. The design architecture included a hairpin aptamer switch probe HP1, consisting of three domains: a DNA aptamer sequence, a caged toehold domain in the stem region blocked by its complementary sequence (toehold hidden), and the 3’ fragment with a ferrocene (Fc) reporter that is capable of acting as a branch migration domain in the subsequent toehold-mediated strand displacement (TMSD) reaction. The biosensor has another component called the I-DNA, which is made to be partially double-stranded by a partially complementary DNA fragment, a blocker DNA. ATP binding triggers TMSD; the branch migration domain displaces the blocker DNA, positioning the HP1 probe, with the Fc at the 3’ end closer to the electrode, generating the electrochemical response.
The biosensor achieved ATP detection as low as 5 nM. A correlation coefficient value is 0.9964 for the 5 nM to 10 nM range. They confirmed the selectivity for ATP by testing other similar molecules: GTP, CTP, UTP, and TTP. They demonstrated ATP-induced liberation of the hidden toehold, subsequently branch migration reaction, generating an electrochemical readout specific to ATP.
Innovation.
FRIH, the human ferritin heavy chain is a large protein, 474 kDa, and is expected to be present in ISF at lower concentrations than in blood. I approached the biosensor design, achieving high sensitivity with the sandwich ELISA concept, a hybrid molecular design involving a DNA aptamer and a nabobody.
Innovation in the design of a caged toehold conjugated to Nb72
Strong nanobody binders such as Nb72, for FRIH, have been reported in the literature (Hu et al. 2022.). I’ve designed an extended ssDNA arm to be conjugated to Nb72. The extended arm includes a hidden toehold and a complement of an invader strand capable of responding to target-induced structural change and subsequently participating in a branch migration reaction.
Innovation in the DNA aptamer design for FRIH:
There is no DNA aptamer for FRIH reported in the literature. This is a gap in the design of a biosensor for ferritin. I am using the computational tools to design a DNA aptamer by screening a randomly generated in silico oligo library for minimal free energy and predicted secondary structures. I am validating the candidate DNA aptamers docking to FRIH in HADDOCK 2.4 modeling.
Innovation in the cell-free synthesis of FRIH.
Ferritin is a 24-mer cage protein. The E. coli expression system has been used for the synthesis and purification of ferritin. But, not from a cell-free protein synthesis reaction. I am proposing to use a cell-free system to obtain purified protein with 24-mer assembly. If it works, this approach is rapid and scalable, allowing for a continuous source of purified protein supply. Oligomeric states of the purified FRIH will be assessed by CDMS analysis using a Waters Xevo instrument.
Innovation in Proximity Ligation Assay (PLA) for de-risking molecular design
PLA is a robust, high-throughput assay, and it can be used to validate the proximity of binders on FRIH to assess whether the binders are in close proximity before microneedle fabrication. The assay is performed in solution with a qPCR readout, translating protein binding to nucleic acid amplification.
Innovation in computational modeling of Nb72.
There is no structural model available for Nb72. Nb72 CDR has been reported, CDR1: IRWSGTST; CDR2: GHTFSSAD; CDR3: AAACDDGLIIRTTVSY (Hu et al. 2022.)I am generating a model for Nb72 by using a common immunoglobulin fold, such as the anti-GFP nanobody (PDB: 3OGO) as a scaffold, grafting Nb72 CDR, and modeling by AlphaFold2. I am using the Nb72 structure model for docking with FRIH by AlphaFold3. Modeling allows for mapping spatial geometry in the complex, and the information can be used for rational epitope exclusion for the design of FRIH DNA aptamers.
Significance and Impact.
This project aims to develop a minimally invasive ISF biosensor for human ferritin. Ferritin is the body’s major iron storage protein. It is the biomarker for iron deficiency. Iron is a nutrient, and yet iron deficiency is prevalent worldwide, affecting people globally, most notably pregnant women, infants, and adolescents. Populations in low- and middle-income countries are experiencing more iron deficiency incidents. Even though incidents of iron deficiency are high, there are only blood-based assays available for testing, requiring lab visits, blood collection, and processing. It can be problematic for rural communities that are not close to hospitals and clinics to have access to laboratory tests. An overload of iron is equally important: an indication of inflammation, infection, liver disease, or malignancy. While ferritin is such an important biomarker, there is no biosensor for ferritin currently available. This project addresses this gap, developing a first-time minimally invasive method that is an off-the-shelf home-use biosensor for ferritin detection.
The most critical barrier for the development of the ISF ferritin biosensor is the size of ferritin. Having a mass of 474 kDa, the size imposes diffusion limitations from serum to ISF. So, the protein is expected to be present at lower concentrations in ISF, as the study demonstrated with other large proteins (Sprunger et al. 2025.) It is necessary to have a highly sensitive detection method for ferritin detection in ISF. An aptamer alone cannot achieve the required sensitivity, and a hybrid modality is essential: a capture nanobody and a reporter DNA aptamer pair.
The second most critical barrier is that there is no DNA aptamer for ferritin ever described in the literature. The success of hybrid modality ISF ferritin detection with proximity-triggered electrochemical readout depends on a DNA aptamer binder and a hybrid nanobody. This project addresses this gap, for the first time generating a computationally designed DNA aptamer and a hybrid nanobody, utilizing the existing nanobody binder Nb72.
Broader implications of this project at the societal level, once the validation is achieved: minimally invasive ferritin monitoring at home could eliminate the expense associated with lab tests and operations and enable large population-level screens and access to health data through early detection before more problematic outcomes, such as anemia development.
The project has technical advancements: a first-time DNA aptamer for ferritin will be identified, adding new tools for ferritin biosensor development. A first-time hybrid nanobody with a caged toehold as a probe will be developed for a molecular architecture that generates electrochemical output through toehold-mediated strand invasion, in which the concept is scalable for other important biomarkers. First-time cell-free expression of a 24-mer protein will be tested. If it confirms that the expected oligomeric state is achieved under cell-free reaction conditions, it simplifies and accelerates protein biosensor development workflow.
The proposed project is the proof-of-concept of a large protein detection from ISF without an enzyme catalyst. If it is achieved, it can be transformative for home-use diagnostic devices and the workflow to develop them. The proposed proximity-ligation assay (PLA) is to de-risk the molecular design before device fabrication. If the PLA workflow is validated, it can be used for the development of other biosensors, such as inflammation-related proteins and NT-proBNP, for cardiac-failure biomarker detection.
Ethical Implications.
The major ethical implications of this project are around clinical testing involving human volunteers, their recruitment, encountering immune responses, and possible misinterpretation or misuse of health data by the general public. Although this project uses an alternative preclinical testing, animal testing is usually carried out at preclinical stages and has ethical implications as well.
In clinical trials, individuals will be recruited to donate ISF samples collected by the microneedle-based device, and their blood samples will be drawn for a correlation study. The device with a hydrogel will stay on the skin for an extended period of time, allowing sample collection and concentration. This is necessary due to the large size of the analyte, as it would be present at a lower concentration in ISF and the hydrogel, and the time of exposure would allow capture and concentration of the analyte. During sample collection, the immune system could potentially trigger a local inflammatory response and sensitization against the materials present on the biosensor, resulting in a skin rash or a delayed response that shows up as hypersensitivity at a later time. The immune response development may be individual-dependent and may be stronger in individuals with underlying conditions. As health data would be readily available from the home tests, misunderstanding of the results and misuse of health data can also occur because people may not have a sufficient technical background on the technology.
To address the ethical implications outlined above, I will put several measures in place. Recruitment for clinical trials will be conducted under a protocol with reviews and approval by the Institutional Review Board (IRB) or an equivalent review committee before the onset of official clinical trial testing. All participants are required to give their consent in written form before they enroll, informing them about the details of the study, including the duration of microneedle-based sample collection, blood withdrawal, and possible risks in the skin’s inflammatory response. To mitigate the anticipated skin’s inflammatory response, the fabrication of the biosensor materials will be chosen from FDA-approved materials and will be compatible with clinical studies. Material safety studies will be conducted with in vitro cytotoxicity tests before human exposure if they are unknown. Participants will be monitored during and after the testing period by qualified health personnel. Protocols for management of adverse skin reactions will be in place for an emergency response, such as topical treatments and early removal of the device. Participants are free to withdraw from the study at any time without any consequence. All data collected will remain in compliance with HIPAA, protecting patient data and safeguarding it to ensure confidentiality.
To overcome possible misuse of information in the general public, a great level of effort and care should be given in preparing educational information about the diagnostic ferritin biosensor. Communication materials should be made available in several outlets, including in-person and online, should address frequently asked questions in a consumer-friendly manner, and should recommend consultations with a healthcare provider about the interpretation of the results.
To preserve animal welfare and follow the ethical principle of not harming, the preclinical testing will be conducted in an alternative way: an artificial ISF spiked with the purified ferritin protein in a 3D-printed platform mimicking the compositional properties of the skin. The 3D-printed system will be operated at physiologically relevant concentrations to assess the efficacy of the biosensor and its molecular components, with an electrochemical readout.
SECTION 4: EXPERIMENTAL DESIGN, TECHNIQUES, TOOLS, AND TECHNOLOGY
Detailed Experimental Plan.
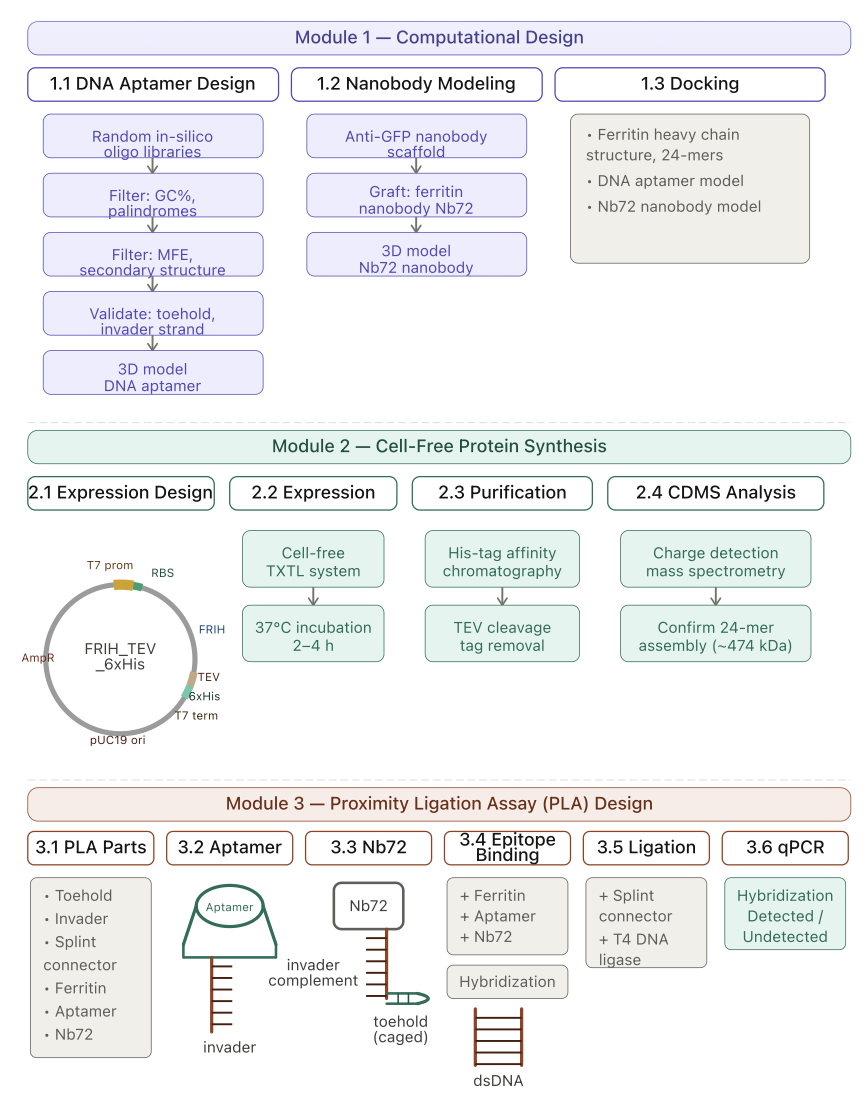
Expected Timeline of the Experimental Plan.
Overview of the Experimental Plan.
This project focuses on the use of computational tools with a rational design and modeling to prioritize the selection of binders to a large protein, FRIH, human ferritin heavy chain, for ISF biosensor development.
SELEX (Systemic Evaluation of Ligands by Exponential Enrichment) has been the gold standard experimental approach for the identification of high-affinity DNA aptamers by screening over many thousands of oligos. Instead of SELEX, I have proposed applying a MAWS workflow (making aptamers without SELEX) by rationally selecting aptamers from in silico DNA oligo libraries. I am applying a filter with criteria known for other binders, such as minimum free energy (MFE) and secondary structure architecture. I am using RNAfold for secondary structure predictions and RNAComposer for 3D structure predictions. I am using HADDOCK 2.4 for DNA aptamer docking to FRIH.
The proposed FRIH biosensor is a dual capture and reporter modality biosensor. DNA aptamers for FRIH are not yet defined in the literature. However, a strong FRIH nanobody binder, Nb72, has been identified. I am using the anti-GFP nanobody structure as a scaffold to model Nb72 by grafting Nb72 CDR onto the scaffold. I am using AlphaFold3 for Nb72 docking to FRIH.
A purified ferritin is needed for the downstream assays to validate binders’ proximity in the proximity ligation assay (PLA), which evaluates the proximity of epitopes bound by the DNA aptamer and the nanobody Nb72.
I have designed a cell-free protein synthesis construct for ferritin expression with a His-tag, allowing purification through a His-tag affinity purification system. A TEV protease cleavage site is also added so that the His-tag can be cleaved off during purification. Ferritin is a 24-subunit oligomeric protein, 474 kDa. I have proposed to determine the oligomeric state of the purified protein by charge detection mass spectroscopy (CDMS) analysis. Cell-free protein synthesis reaction and purification experiments can potentially be performed on cloud labs, and the CDMS analysis can potentially be done at Waters.
PLA is a laboratory assay. Since this project is based on computational design, I focused on the DNA design aspect of PLA, the design of the extended arm sequences, as a concept. NUPACK is widely used in aptamer molecular design. The extended arm sequences are conceptually designed based on published results in the literature. Future work is needed to validate the design by NUPACK. By design, when both binders are in proximity, the extended arms would be responsive to target-induced structure changes and subsequently, participate in the branch migration reaction. PLA requires a splint connector and T4 DNA ligase to be added to seal the nick and circularize the hybridized product, and finally, a qPCR is performed as a readout.
The yeast display assay is a laboratory assay. I’ve proposed this assay to validate DNA aptamer binding to FRIH. Yeast cells are engineered for the expression and surface display of the target protein. Binders are tagged with a fluorescence probe and allowed to interact with the recombinant yeast cells with surface display of the target protein. Yeast cells are passed through flow cytometry, and cells with strong fluorescent signals are sorted and collected, validating DNA aptamer binding to FRIH in an in vivo assay.
Techniques Relevant to This Project.
Bioethical Considerations; DNA Construct Design; Databases (GenBank, NCBI, RCSB); Designing a Twist Order; Protein Design; Use of Benchling; Models and Notebooks; Cell Free Reactions; Protein Purification
Selected Techniques and Their Applications in This Project.
DNA Construct Design.
In the proposed project, FRIH would be purified and added to the proximity ligation assay (PLA) to validate co-binding by the DNA aptamer and the nanobody Nb72. If both binders are in proximity, the aptamer’s invader strand hybridizes to a complementary strand in the Nb72 extended arm to form a double-stranded DNA. The hybridization event can be detected after T4 DNA ligase by qPCR.
To obtain the purified FRIH component, I’ve designed a DNA construct for a cell-free protein synthesis system in which FRIH is transcribed and translated into the ferritin protein.
The amino acid sequence of FRIH, the human ferritin heavy chain, was obtained from the RCSB database and reverse translated to obtain a nucleotide sequence. The DNA construct parts are retrieved from database searches, already existing and well-characterized expression systems: RNA polymerase-dependent promoter, T7 promoter, ribosome binding sequence (RBS), RNA polymerase termination signal, 6xHis tag for his tag affinity purification, and a TEV protease cleavage recognition sequence for removal of the his tag.
Use of Benchling.
DNA parts described above were assembled in silico in Benchling with a plasmid backbone, pUC19. Parts are translated and annotated, ensuring start and stop codons are present on the nucleotide sequence. The use of restriction enzymes in cloning: I ensured that the chosen enzymes are compatible with the cloning procedure. For this, I have screened restriction enzyme recognition sequences in all parts by using the enzyme recognition sequence identification tool in Benchling. The plasmid construct was assembled in silico and will be ordered from Twist Bio as a clonal gene synthesis.
SECTION 5: RESULTS and QUANTITATIVE EXPECTATIONS
This project is based on computational approaches. I’ve focused on two design aspects of this project for validation:
- The DNA construct.
I’ve validated the DNA construct by simulating in Benchling to ensure that cloning of the DNA parts can be achieved as expected when HindIII and SphI restriction digestion are performed on the plasmid DNA (see MODULE 2). Validation by laboratory tests has been planned by ordering the construct from Twist Bio and potentially testing the cell-free synthesis at Ginkgo Cloud Labs (Nebula). Following purification, the oligomerization state of the purified protein is planned to be potentially analyzed by charge state mass spectroscopy (CSMS) at Waters.
- DNA aptamer and nanobody binders and docking.
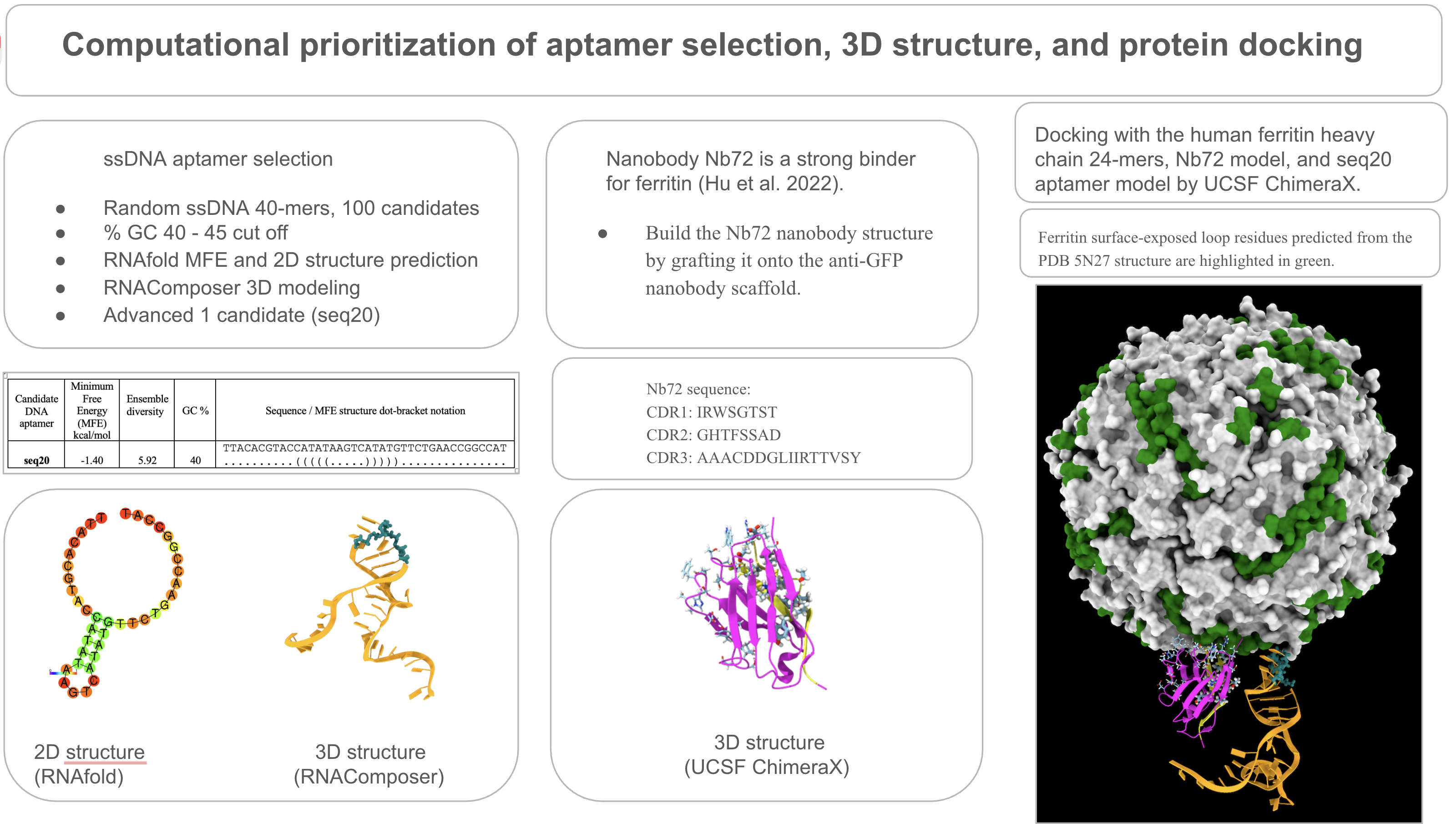
I’ve validated binders by modeling the following: a) the candidate DNA aptamer, Seq 20, and b) nanobody binder, Nb72.
The DNA aptamer Seq 20 was validated by a structure modeling with RNAComposer (see MODULE 1, Structure modeling of aptamer Seq 20). Further validation was obtained by the HADDOCK 2.4 modeling for docking to FRIH (see MODULE 1, Docking: Aptamer Seq 20 to FRIH).
The nanobody Nb72 model was validated by structure modeling by AlphaFold2 (see MODULE 1, Structure modeling of Nb72). Further validation was obtained by the AlphaFold3 modeling for docking to FRIH (see MODULE 1, Docking: Nb72 to FRIH ).
MODULE 1
Randomly generate 40-mer ssDNA in-silico library.
I’ve used the computational tool, bioinformatics.org, to generate a 100-random ssDNA in silico library.
GC % filter and palindromic exclusion.
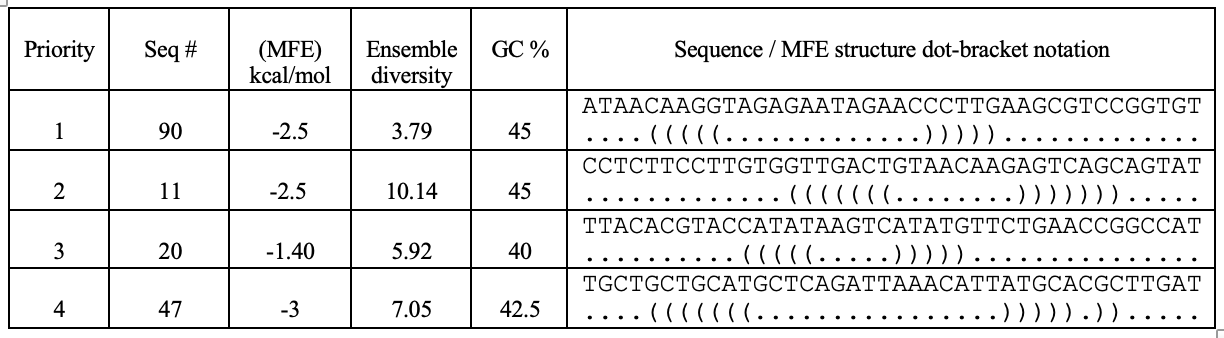
GC % filtering and exclusion of palindromic repetitiveness yielded 12 priority candidates. The GC % cut-off was between 40 - 45 %, based on the established design principles of aptamer binders.
| Seq # | GC % | sequence |
|---|---|---|
| 20 | 40.0 | TTACACGTACCATATAAGTCATATGTTCTGAACCGGCCAT |
| 41 | 40.0 | CATTAATATGGTTGAGGCGGACTTTATACATCGACTCGTA |
| 33 | 40.0 | TATCCTAAGTAACTAACAGCAGATCAGCAAAGAGAGATGG |
| 36 | 40.0 | CAATGTCCTAGAGTAATTGTGAACACTTTAAGCTGCTGAC |
| 97 | 40.0 | GCGTACTTGCAACGAGGCTGATTAACATGTCTAATGAATA |
| 10 | 42.5 | TAGTATGGTCGGAGAAGTTAATGATGGTTTAGCCGTGTCA |
| 47 | 42.5 | TGCTGCTGCATGCTCAGATTAAACATTATGCACGCTTGAT |
| 80 | 42.5 | TAGATTTGAAGTTACCGTGTTGCCTGACCGTAACTTCTGA |
| 5 | 45 | ATCGGCTTGAAATCTGATGCTCTTAAAGTGTCTGCAGGCT |
| 11 | 45 | CCTCTTCCTTGTGGTTGACTGTAACAAGAGTCAGCAGTAT |
| 53 | 45 | AGTCTTATAACTGGCGAGCTCGAAGGAAGTACGTTGAATC |
| 90 | 45 | ATAACAAGGTAGAGAATAGAACCCTTGAAGCGTCCGGTGT |
Secondary structure filtering.
I’ve used RNAfold (Vienna RNA Web Server) to filter predicted secondary structures based on their minimum free energy (MFE) and secondary structure folds.
RNAfold parameters selected.
minimum free energy (MFE) and partition function
avoid isolated base pairs
Incorporate G–Quadruplex formation into the structure prediction algorithm
dangling energies on both sides of a helix in any case
DNA parameters (Matthews model, 2004)
MFE.
The candidate should have a moderate MFE, ensuring fold stability and accommodating toehold opening. As a first pass, I used the MFE values close to between -3 and -10 kcal/mol.
Secondary Structure.
Prioritize a candidate binder with a defined stem-loop structure. DNA aptamers are known to make contact through exposed loops that bulge out or at junctions. Screen candidates that have clean, accessible stem-loop architecture.
The end of the structure should not be paired; it should be free to accept an arm extension. The ends should have dots in the dot bracket notation, indicating that the ends are free.
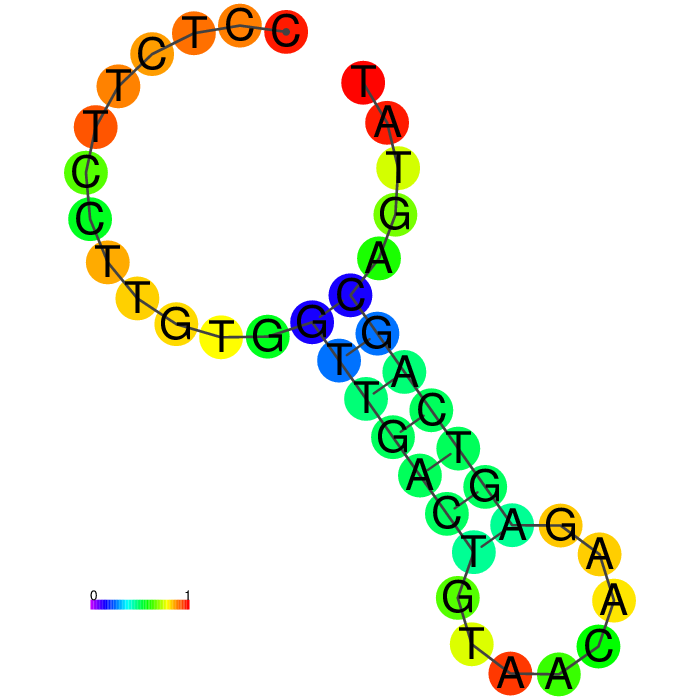
The Heatmap.
The loop residues are red and orange, indicating they are likely to be unpaired within the structure and are likely to be accessible to make contact with the protein. Blue and cyan residues are likely to be paired.
Ensemble diversity.
Prediction for how many confirmations could occur. A low number indicates a well-defined, consistent structure, and the structure is more likely to remain in the same conformation.
A summary table showing secondary structure analysis of 12 DNA aptamer candidates.

Figure showing predicted secondary structures of the candidate aptamers.
| Seq 20 | Seq 41 | Seq 33 | Seq 36 |
|---|---|---|---|
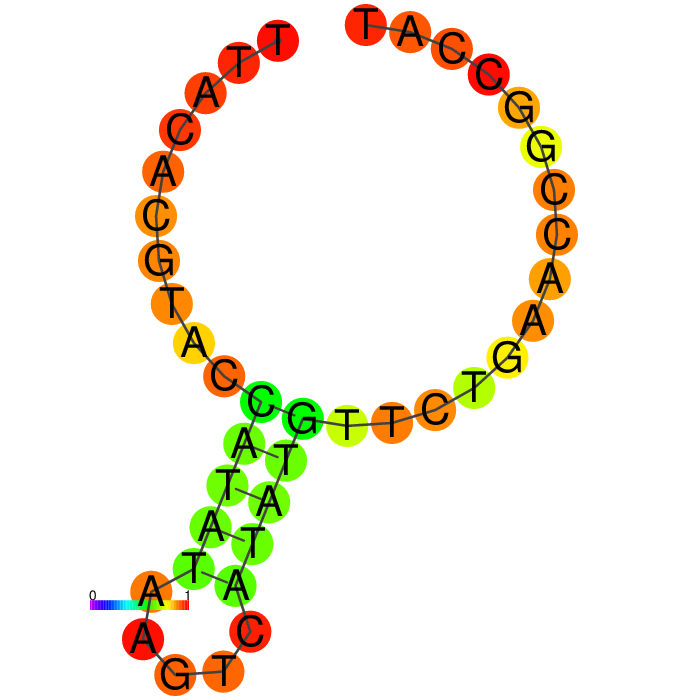
| 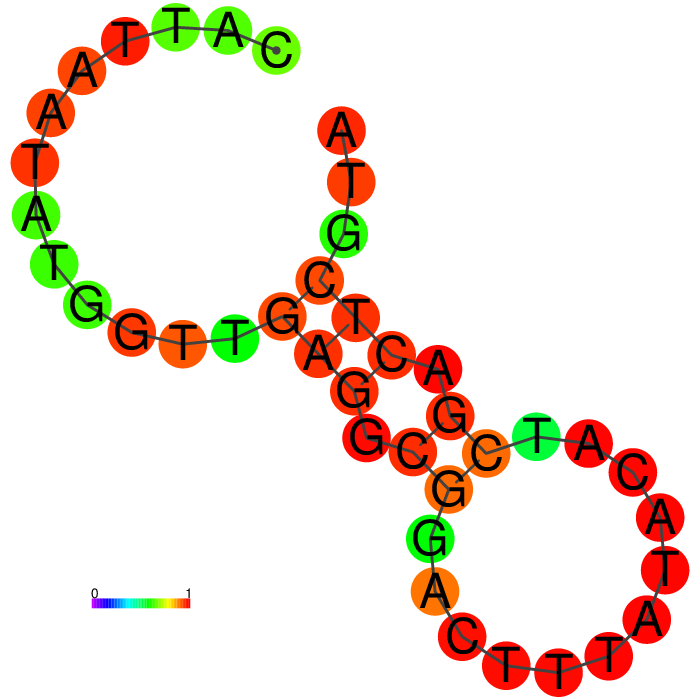
| 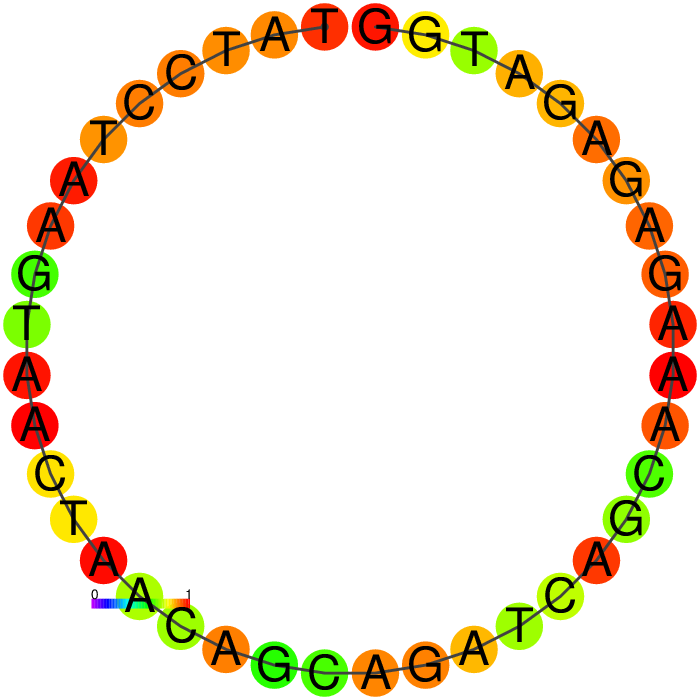
| 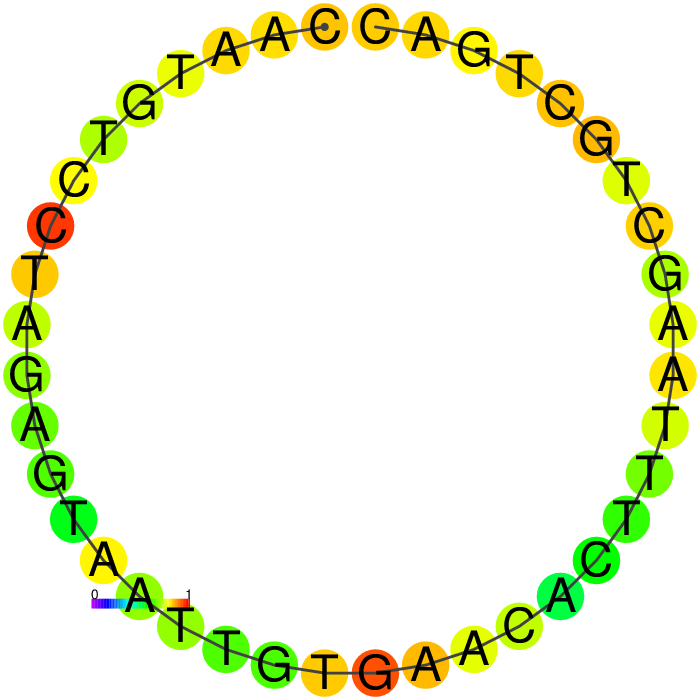
|
| Seq 97 | Seq 10 | Seq 47 | Seq 80 |
|---|---|---|---|
| 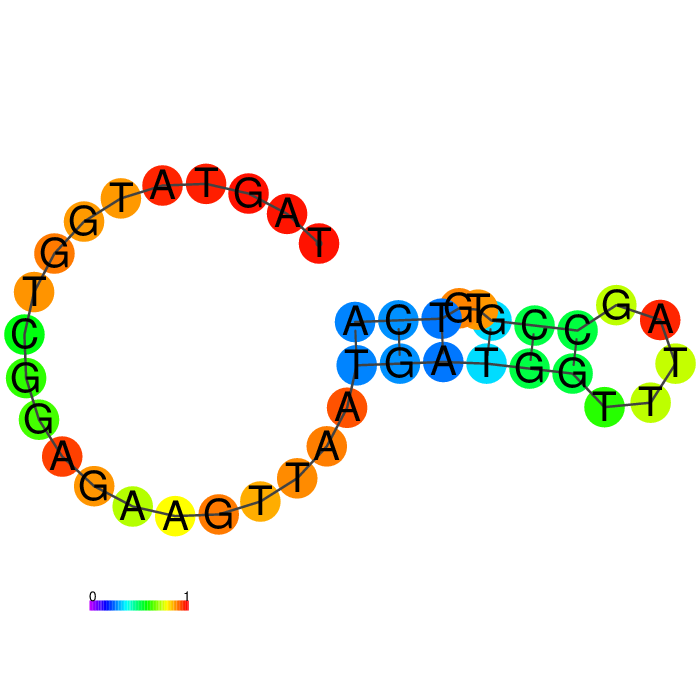
| 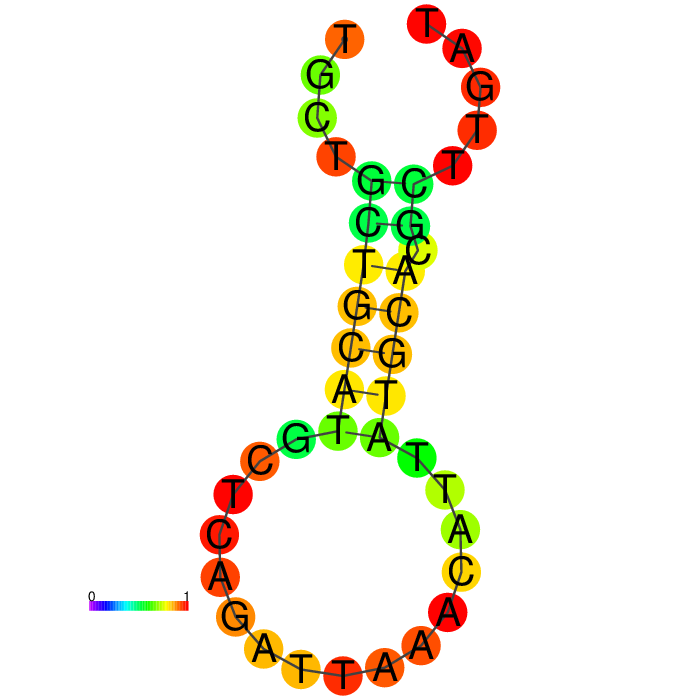
| 
|
| Seq 5 | Seq 11 | Seq 53 | Seq 90 |
|---|---|---|---|
| 
| 
| 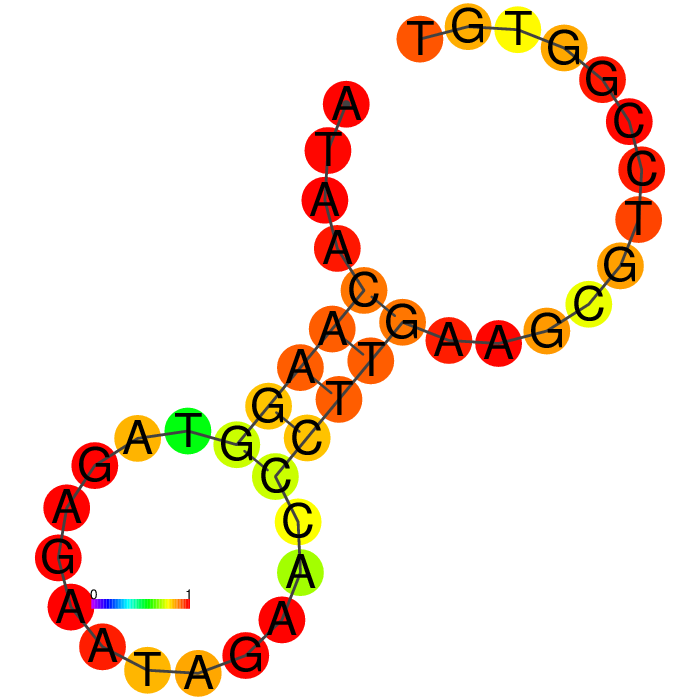
|
Base-pairing probability color key
The top four candidates were selected and prioritized based on their MFE and folding with clean, accessible stem-loop architecture.
Table summarizing secondary structure analysis of the top four candidates.
Validating secondary stem-loop structure with an extended arm sequence.
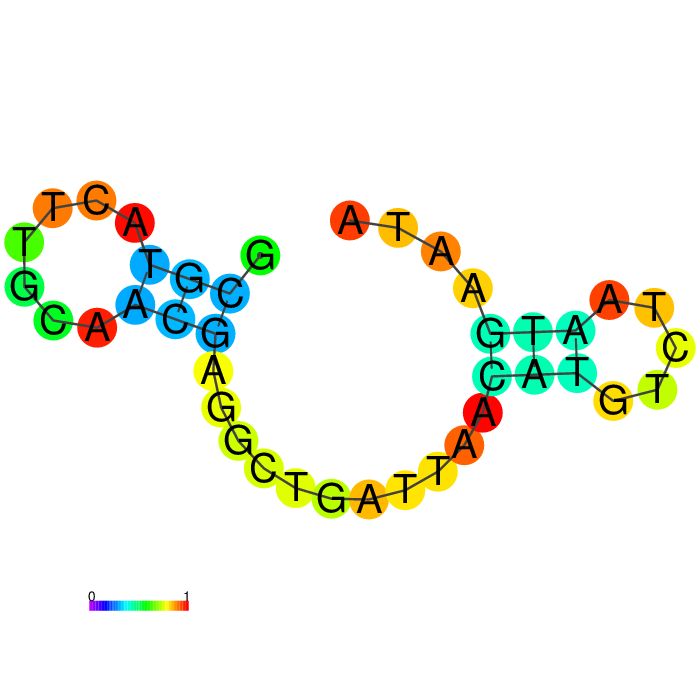
The proposed FRIH biosensor is a dual modality sensor with capture and reporter binders, uses a toehold-mediated strand invasion mechanism (TMSI). Aptamer design is responsive to target-induced structure changes, generating an electrochemical output. The aptamer has an extended arm consisting of two domains, an invader strand and a toehold complementary sequence. The capture nanobody has an extended arm with a caged toehold (hidden) and a complement to the invader strand. Because an extended arm sequence may affect the aptamer's binding to the target, the aptamer's secondary structure should be evaluated with the prospective extended arm.
Wang et al. 2015 paper describes a closely related technology, a toehold-mediated strand displacement (TMSD) mechanism with a caged toehold responsive to target binding. I’ve adopted the idea that FRIH binding triggers the opening of the caged toehold, subsequently branch migration reaction by the invader strand. I’ve applied design principles for caged toehold and TMSD to create the domains of the extended arm. (Wang et al. 2015 and Zhang and Winfree 2009), and evaluated those aptamers for 2D structure stability for validation.
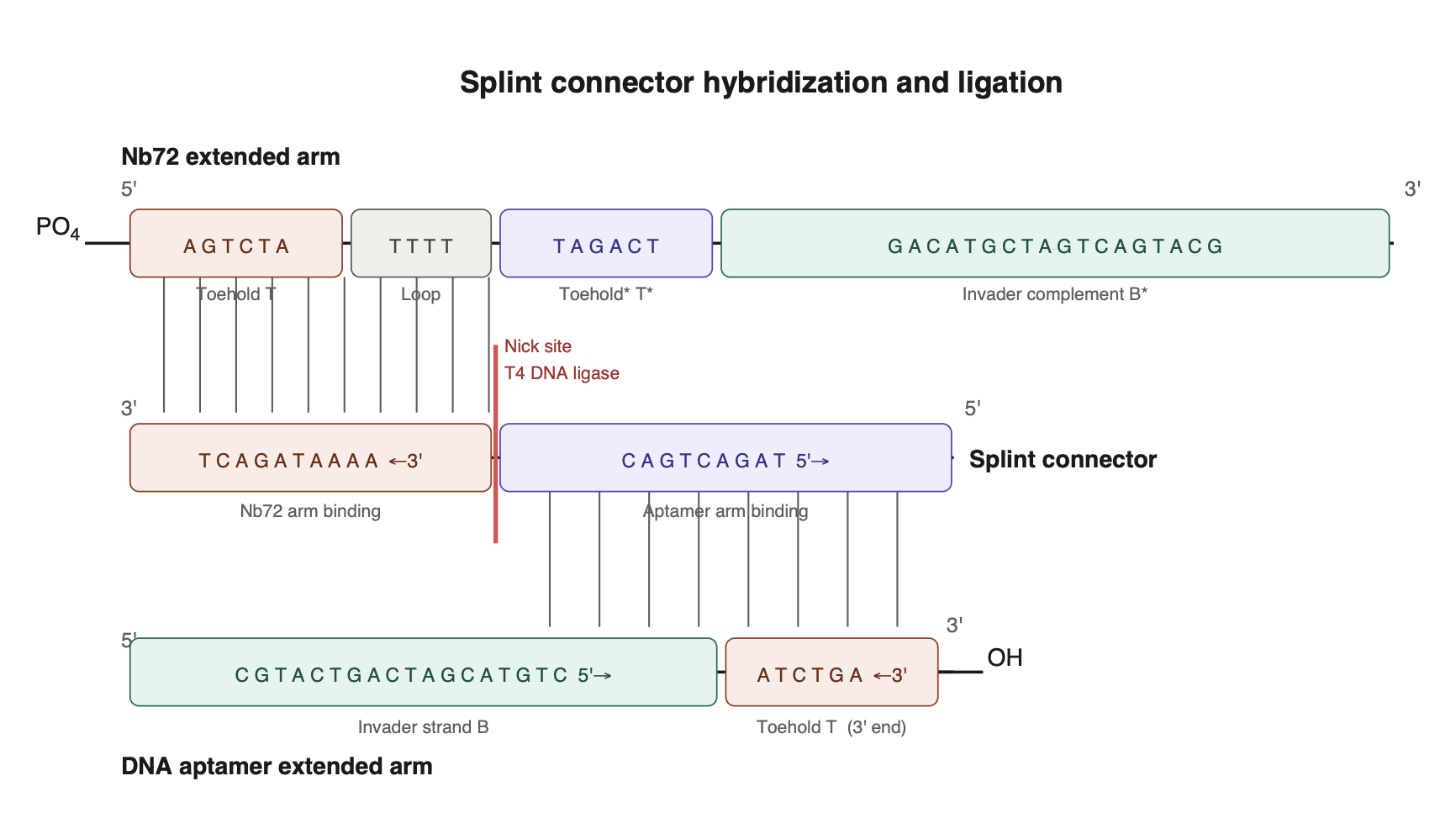
The extended arm architecture includes: a 6-nt toehold complement, an 18-nt invader strand, and a 4-nt linker, which provides flexibility in the junction.
Tables showing the domains of the extended arms.
| Domains in aptamer extended arm | Sequence 5’ - 3’ |
|---|---|
| Toehold complement - T* | TAGACT |
| Invader strand - B | CGTACTGACTAGCATGTG |
| Linker | TTTT |
| Domains in Nb72 extended arm | Sequence 5’ - 3’ |
|---|---|
| Toehold complement – T* (caged) | TAGACT |
| Loop | TTTT |
| Toehold – T | AGTCTA |
| Invader strand complement – B* | GACATGCTAGTCAGTACG |
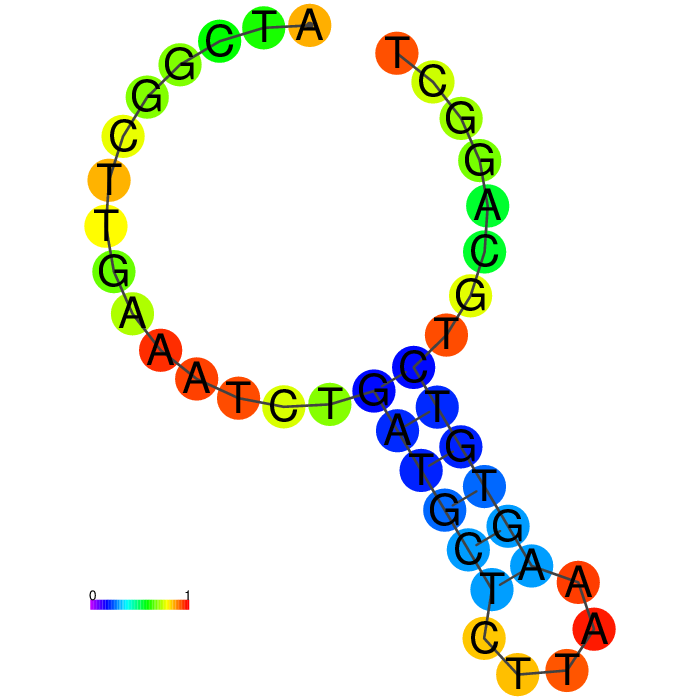
Figure showing secondary structures of aptamers and the aptamers with extended arms.
| Seq 90 aptamer | aptamer-extended | aptamer-extended plain structure |
|---|---|---|
|
| 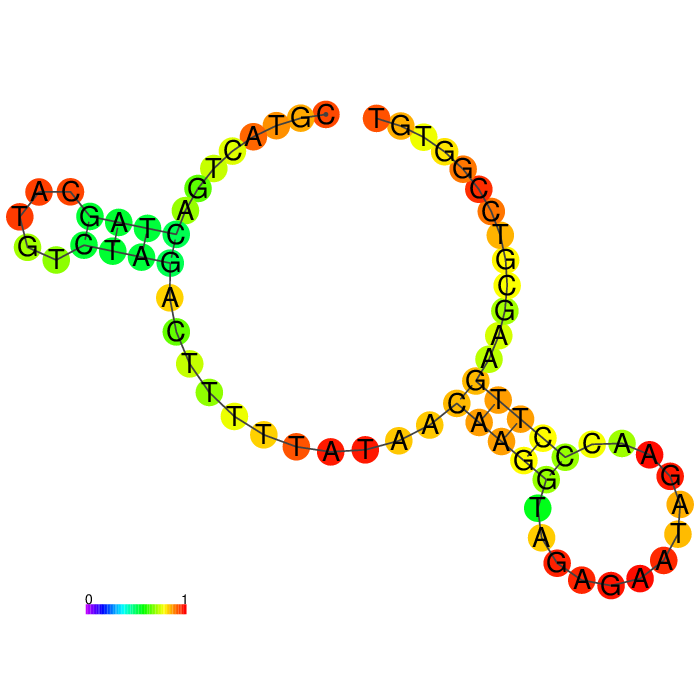
| 
|
| Seq 11 aptamer | aptamer-extended | aptamer-extended plain structure | |
|---|---|---|---|
|
| 
| 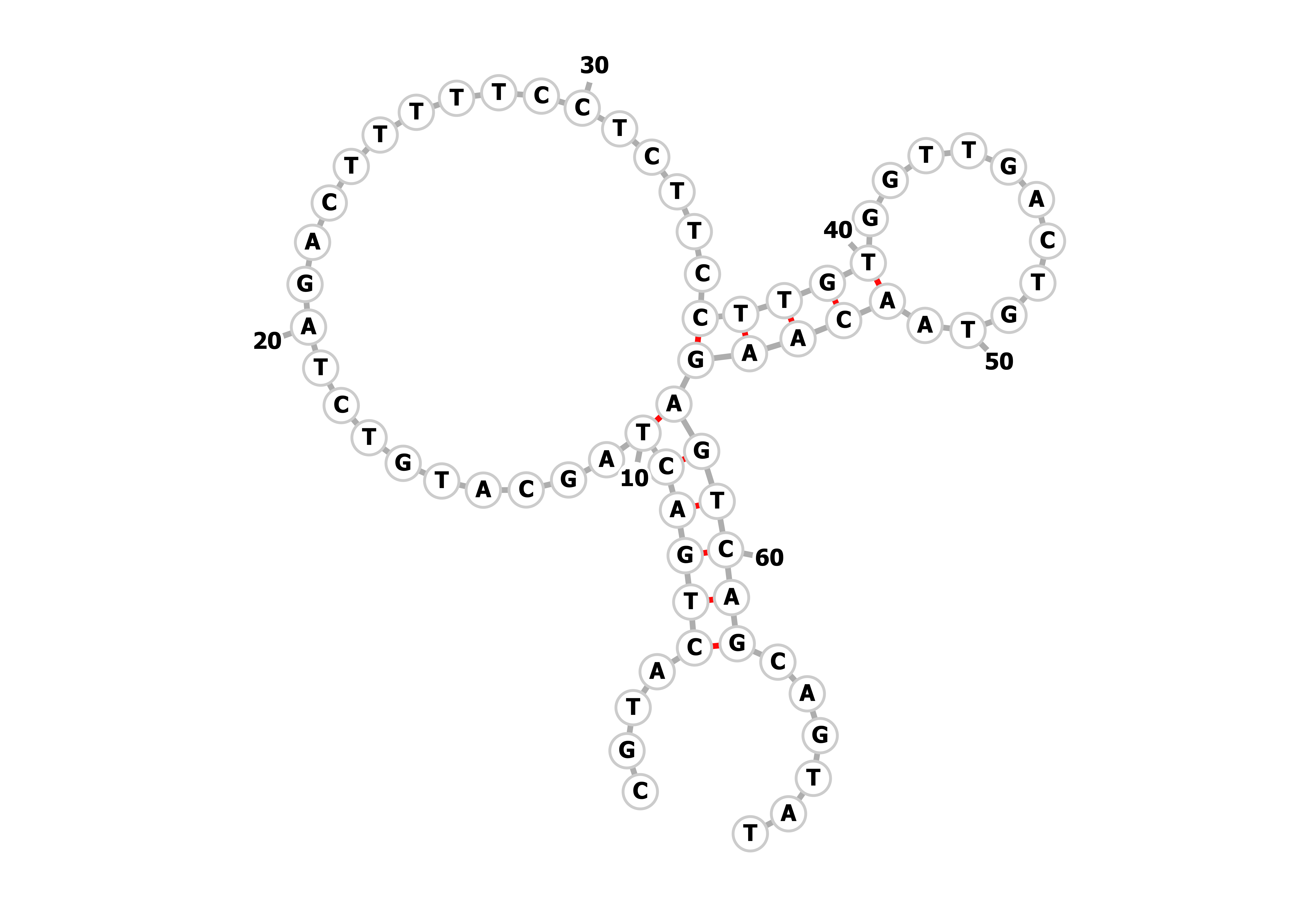
|
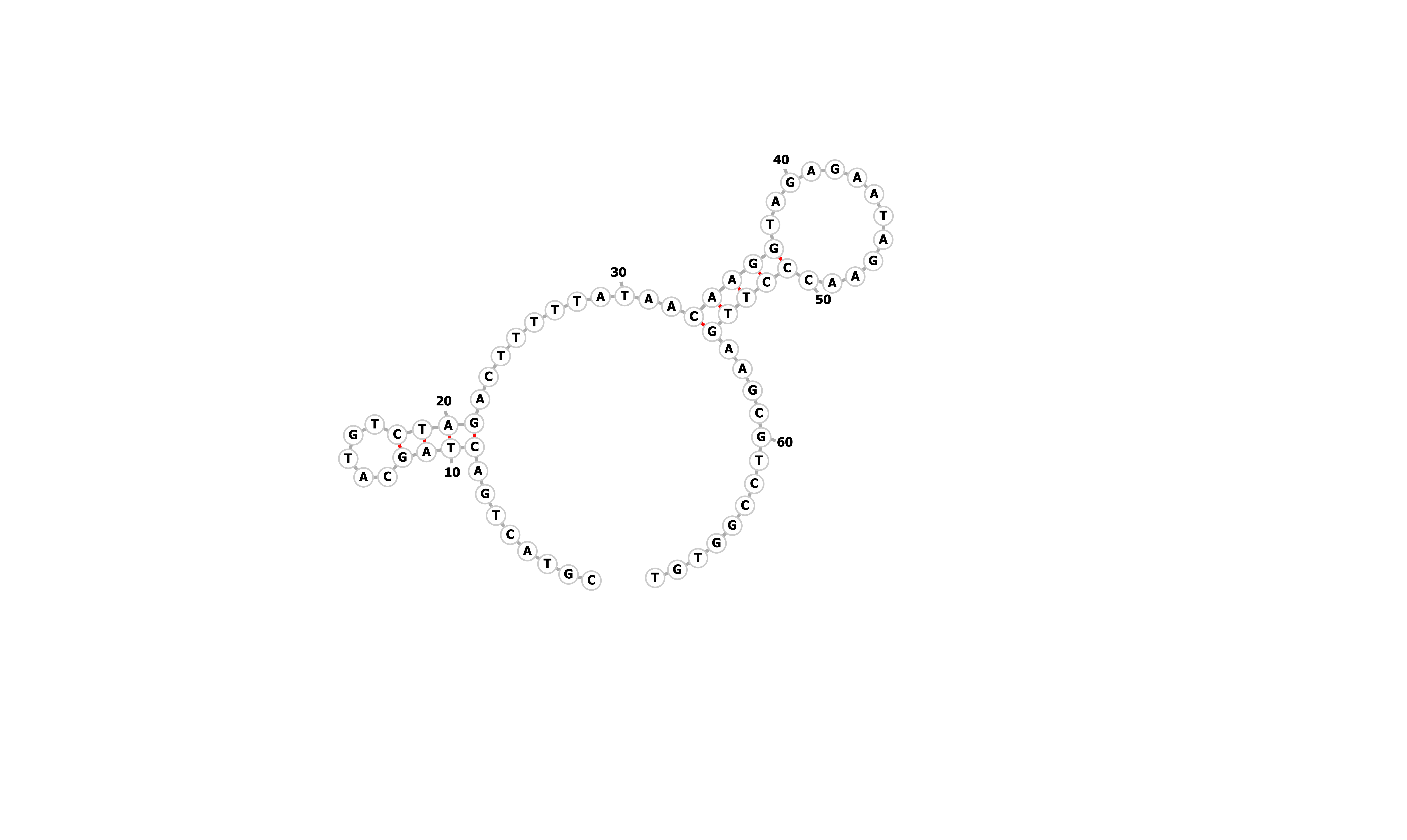
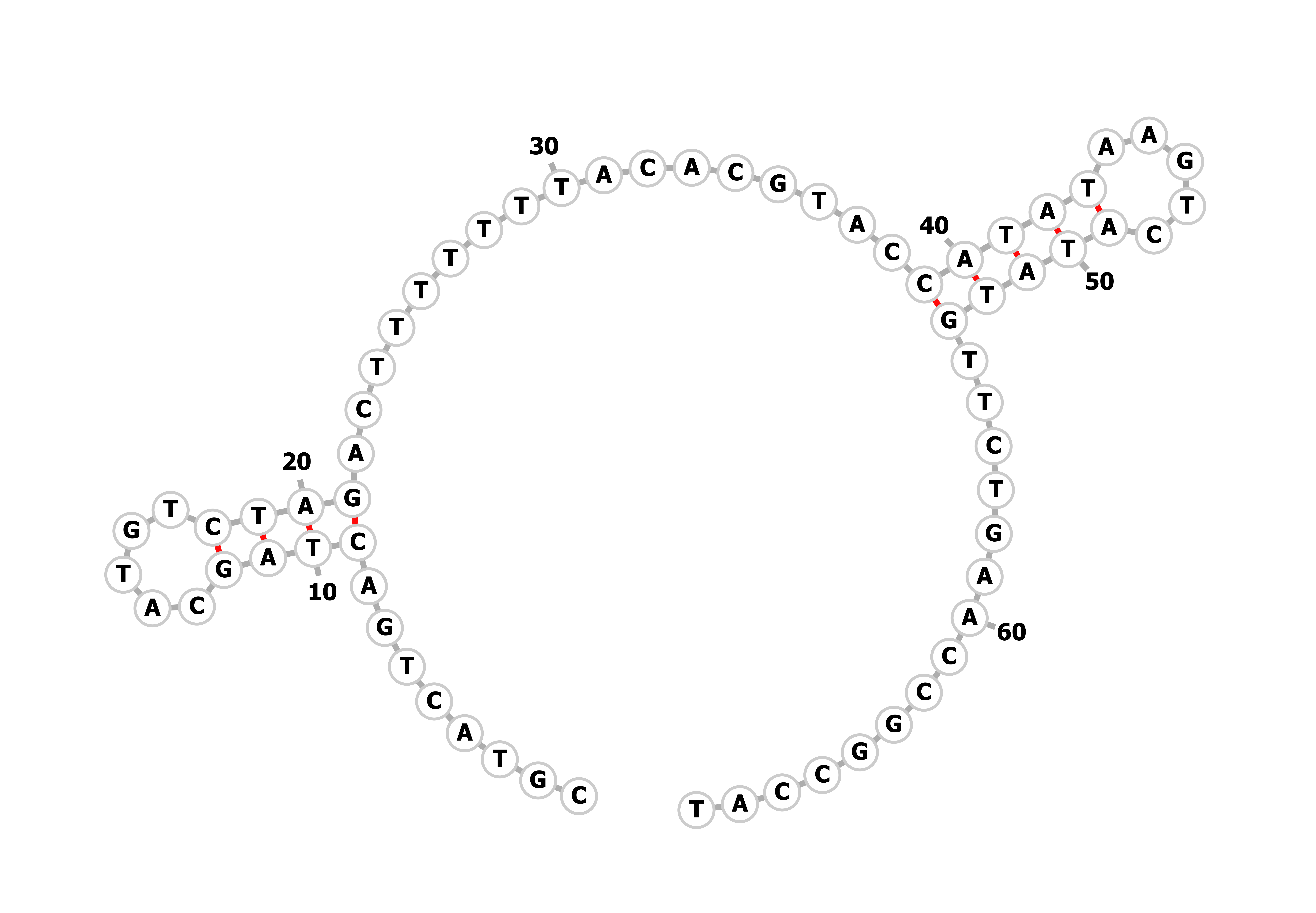
| Seq 20 aptamer | aptamer-extended | aptamer-extended plain structure | |
|---|---|---|---|
|
| 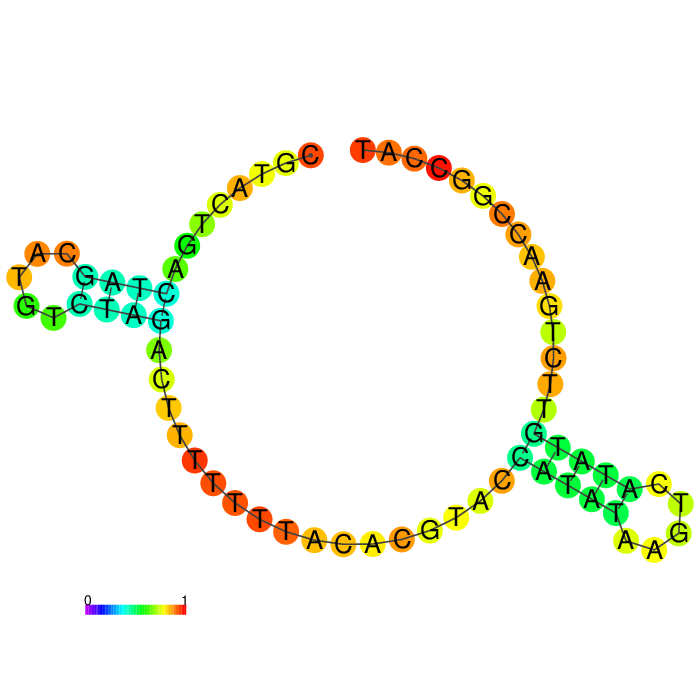
| 
|
| Seq 47 aptamer | aptamer-extended | aptamer-extended plain structure | |
|---|---|---|---|
|
| 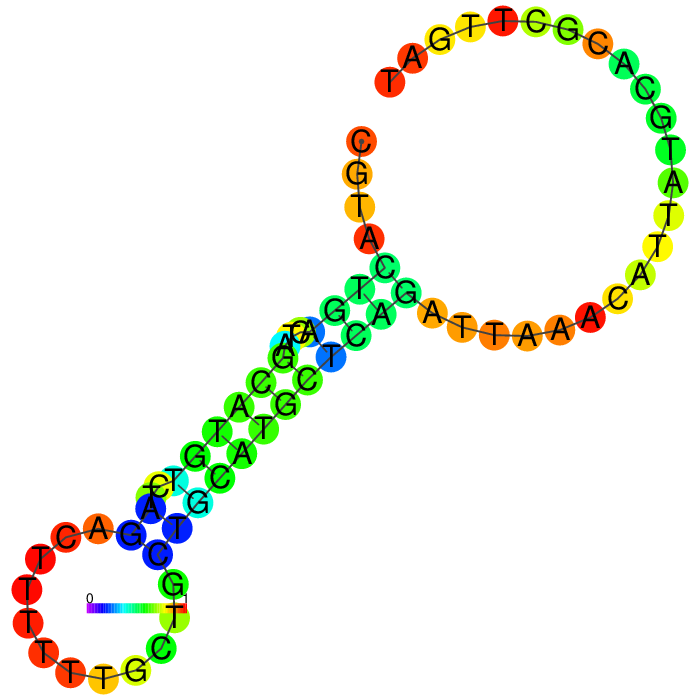
| 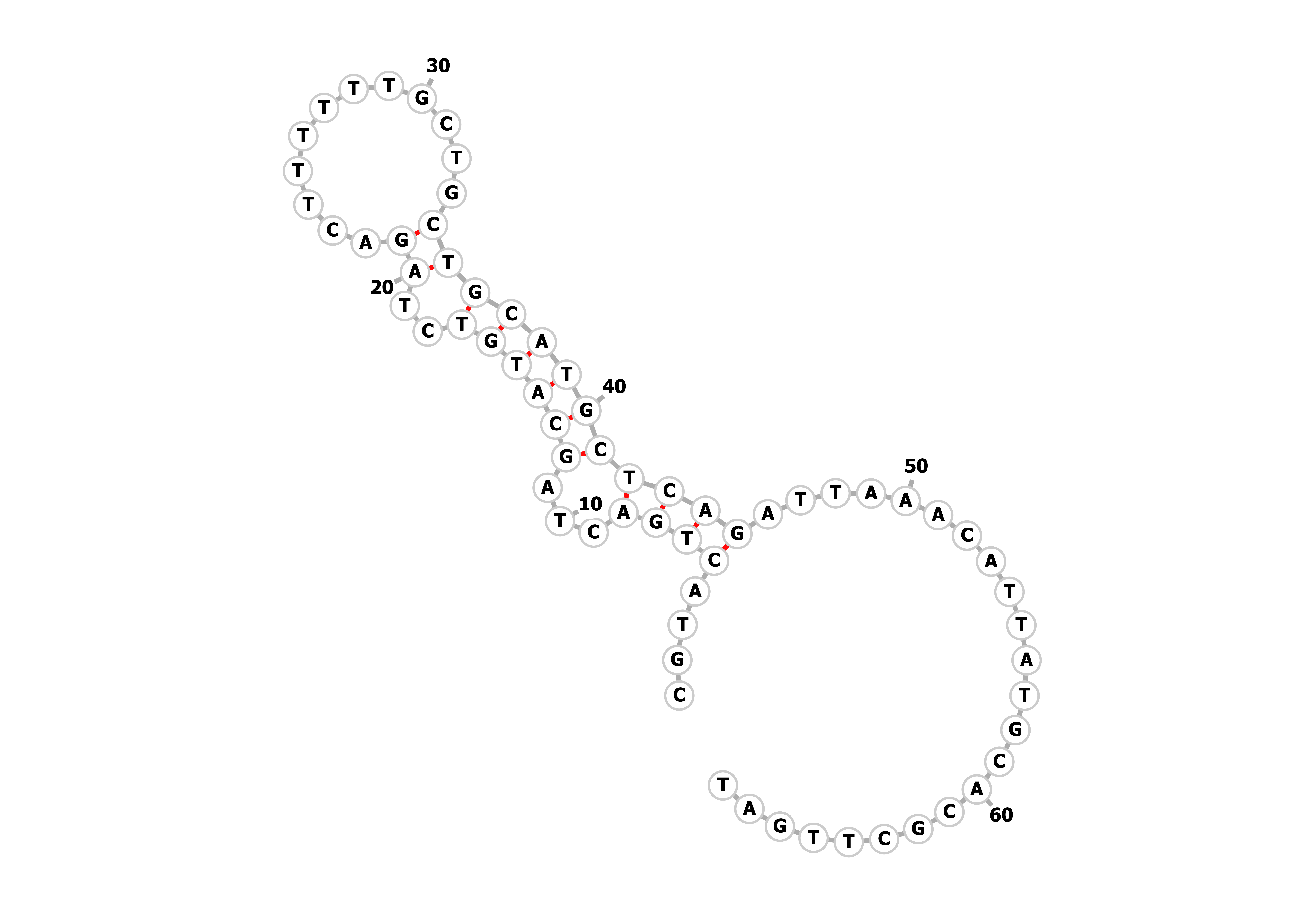
|
Base-pairing probability color key
Table summarizing secondary structures of the aptamers and aptamers with extended arms.
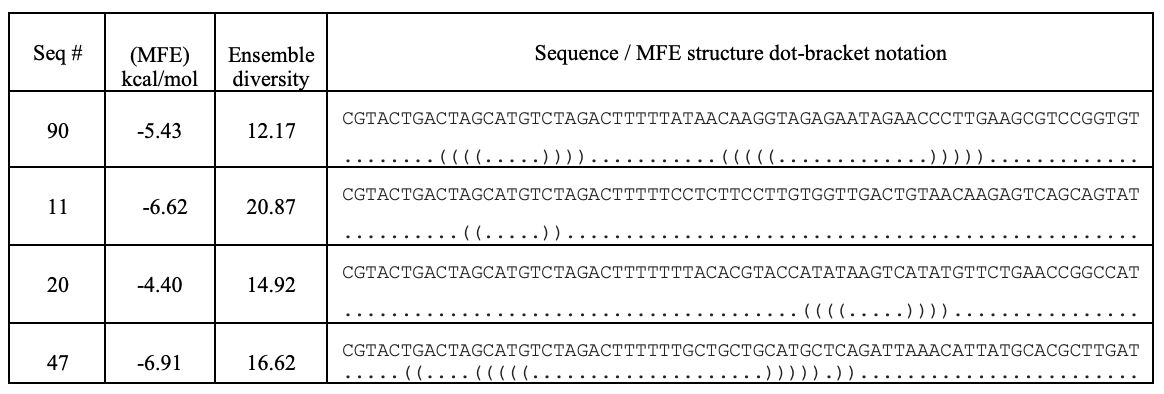
Aptamer Seq 20 was advanced to be the top DNA aptamer candidate.
Aptamer Seq 20 with an extended arm had a clean stem-loop architecture with moderate MFE. The rest of the candidates had a significant change in stem-loop architecture. Therefore, they are de-prioritized.
Table showing predicted solvent-exposed (unpaired) residues of aptamer Seq 20.
| Position | Residue | Region | Pairing status | Priority for docking |
|---|---|---|---|---|
| 1-10 | TTACACGTAC | 5’ tail | Unpaired | Medium |
| 11-15 | CATAT | Stem | Paired | Exclude |
| 16-20 | AAGTC | Hairpin loop tip | Unpaired | High |
| 21-25 | ATATG | Stem | Paired | Exclude |
| 26-30 | TTCTG | Stem-loop junction | Unpaired | High |
| 31-40 | AACCGGCCAT | 3’ tail | Unpaired, fold back | Low |
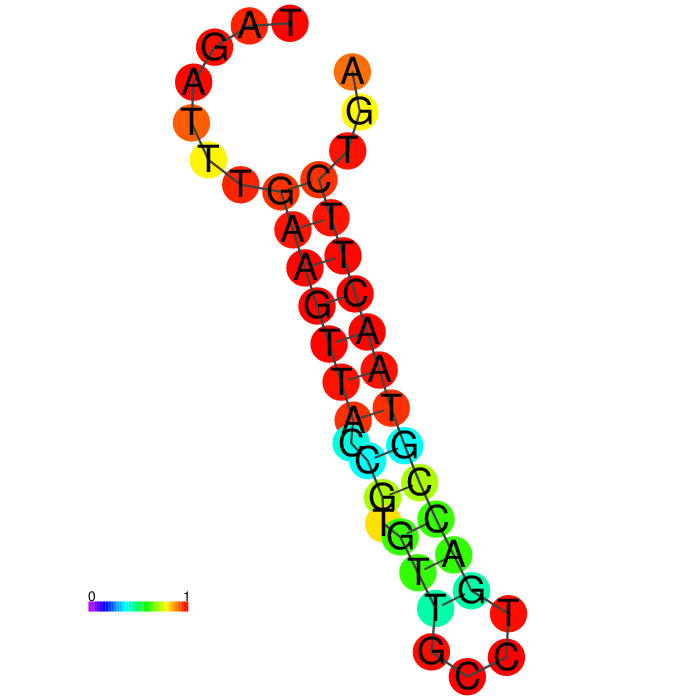
Structure modeling of aptamer Seq 20.
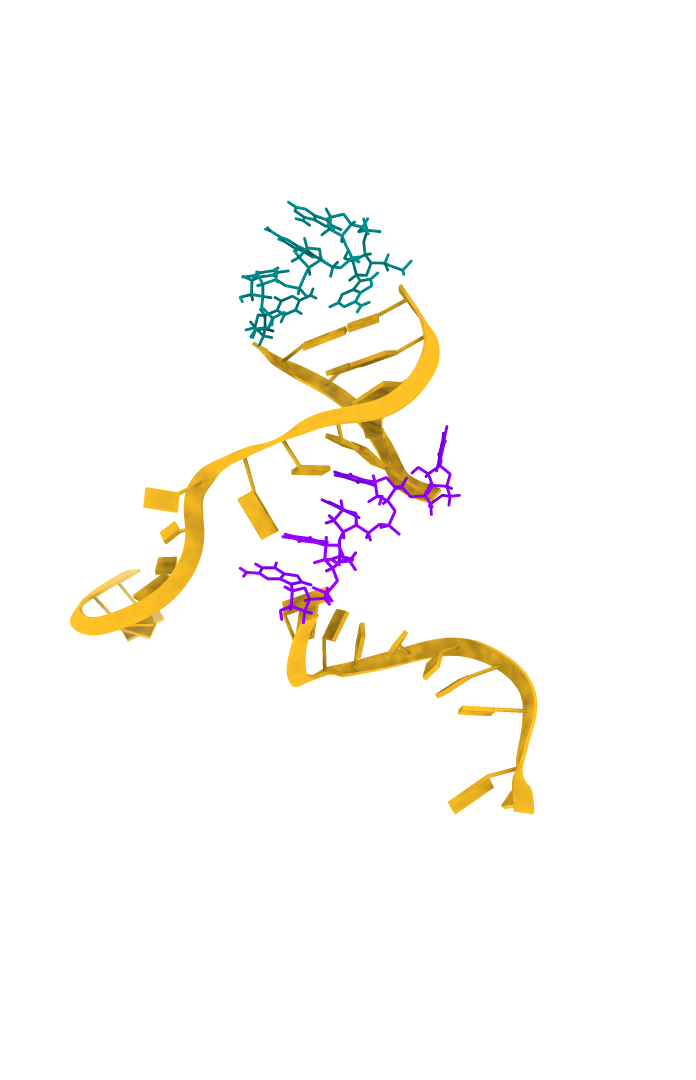
Below are the 3D models of the DNA aptamer Seq 20 generated by RNAComposer and visualized by ChimeraX in a schematic nucleotide/slab representation. Predicted active residues in the ball-and-stick, the hairpin loop tip, 16-20 (teal), and the stem-loop junction residues, 26-30 (purple).
| DNA Aptamer Seq 20 | Predicted active residues (ball-and-stick) |
|---|---|

| 
|
Structure modeling of Nb72.
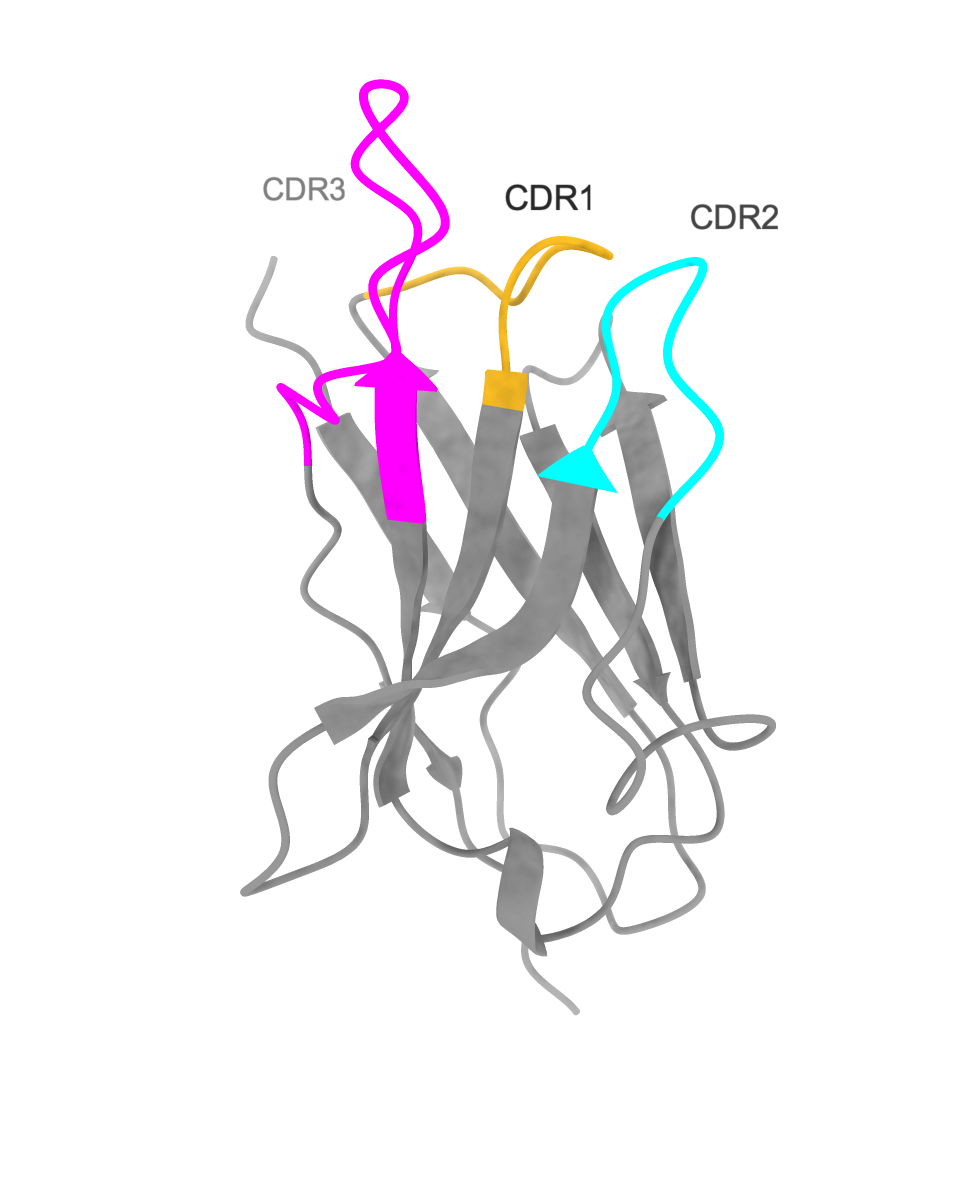
Nanobodies consist of relatively conserved beta sheet scaffolds and variable complementarity-determining regions (CDRs) that bulge out to form loops. The scaffold does not make contact with the antigen, but the variable regions in CDRs do. Hydrophobic residues in CDR1 form stacking interactions with aromatic residues of the antigen, thereby contributing to the positioning of the binding surface. CDR2 has a role in discriminating closely related antigens, achieving specificity. CDR3 is the most important CDR of all; it makes direct contact with the antigen.
Nb72 is a strong nanobody binder of the human ferritin heavy chain, and the Nb72 CDR has been identified by Hu et al. 2022. Although the structure of Nb72 is currently lacking, a structural model can be computationally built based on a conserved immunoglobulin fold, such as the anti-GFP nanobody structure, PDB: 3OGO Kubala et al. 2010.
Table showing Nb72 CDR and amino acid sequence.
| Nb72 | Number of residues | Amino acid |
|---|---|---|
| CDR 1 | 1-8 | IRWSGTST |
| CDR 2 | 1-8 | GHTFSSAD |
| CDR 3 | 1-16 | AAACDDGLIIRTTVSY |
Table showing the anti-GFP nanobody CDR and amino acid sequence of 3OGO chain E.
| Anti-GFP | Residue range | Amino acid |
|---|---|---|
| CDR 1 | 28-34 | FPVNRYS |
| CDR 2 | 52-59 | MSSAGDRS |
| CDR 3 | 98-105 | NVNVGFEY |
Table showing the residue range in the grafted amino acid sequence of 3OGO chain E.
| Nb72 | Residue range | Amino acid |
|---|---|---|
| CDR 1 | 28-35 | IRWSGTST |
| CDR 2 | 53-60 | GHTFSSAD |
| CDR 3 | 99-114 | AAACDDGLIIRTTVSY |
- Nb72 modeling by AlphaFold2
To build the Nb72 nanobody model, firstly, I’ve generated an anti-GFP nanobody scaffold based on the 3OGO structure, retrieved from RCSB, anti-GFP nanobody chain E. The next step was to mutate and graft Nb72 on the anti-GFP nanobody chain E framework. Because the CDR3 of Nb72 is longer than that of the anti-GFP nanobody, a simple residue swap would not generate an accurate grafting and modeling. So, I ran the entire grafted amino acid sequence on AlphaFold2 on Colab (ColabFold v1.6.1: AlphaFold2 using MMseq2), which would handle the longer CDR3 by loop engineering. As a reference, I’ve also obtained the anti-GFP nanobody model by AlphaFold2.
The anti-GFP nanobody amino acid sequence, highlighting CDR residues.
MQVQLVESGGALVQPGGSLRLSCAASGFPVNRYSMRWYRQAPGKEREWVAGMSSAGDRSSYEDSVKGRFTISRDDARNTVYLQMNSLKPEDTAVYYCNVNVGFEYWGQGTQVTVSSK
The grafted amino acid sequence, highlighting Nb72 CDR.
MQVQLVESGGALVQPGGSLRLSCAASGIRWSGTSTMRWYRQAPGKEREWVAGGHTFSSADSYEDSVKGRFTISRDDARNTVYLQMNSLKPEDTAVYYCAAACDDGLIIRTTVSYWGQGTQVTVSS
AlphaFold2 parameters selected.
template-mode: none num_relax: 1 msa_mode: mmseq2_uniref_env paired_mode: unpaired_paired model_type: auto num_recycles; 3 template_ recycle_early_stop_tolerance: auto relax_max_iterations: 200 pairing_strategy: greedy max_msa: auto num_seeds: 1
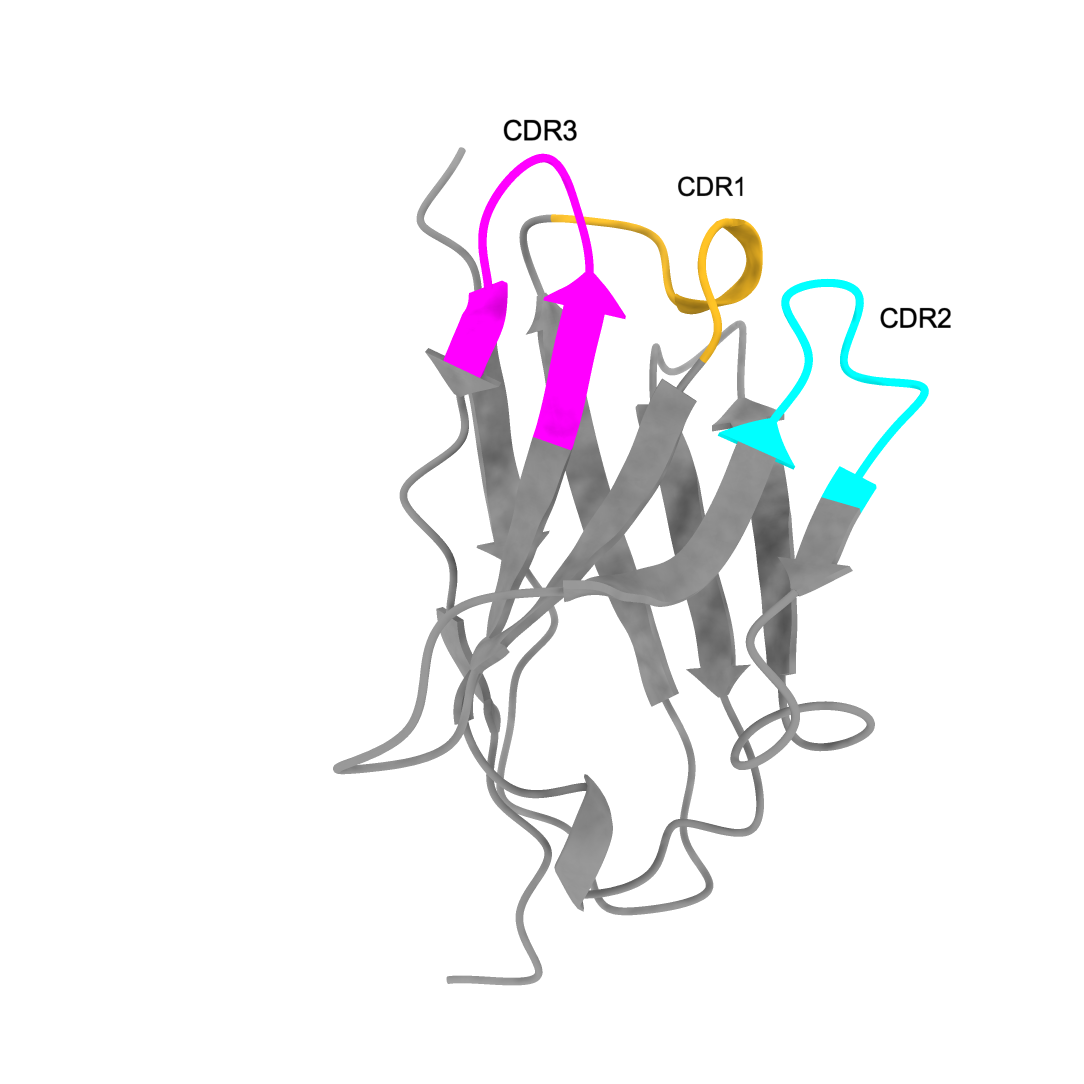
The anti-GFP nanobody model by AlphaFold2 visualized by ChimeraX. pLDDT: 96.9, pTM: 0.902.
The Nb72 nanobody model by AlphaFold2 visualized by ChimeraX. pLDDT: 87.9, pTM: 0.854.
| Anti-GFP nanobody | CDR highlighted |
|---|---|
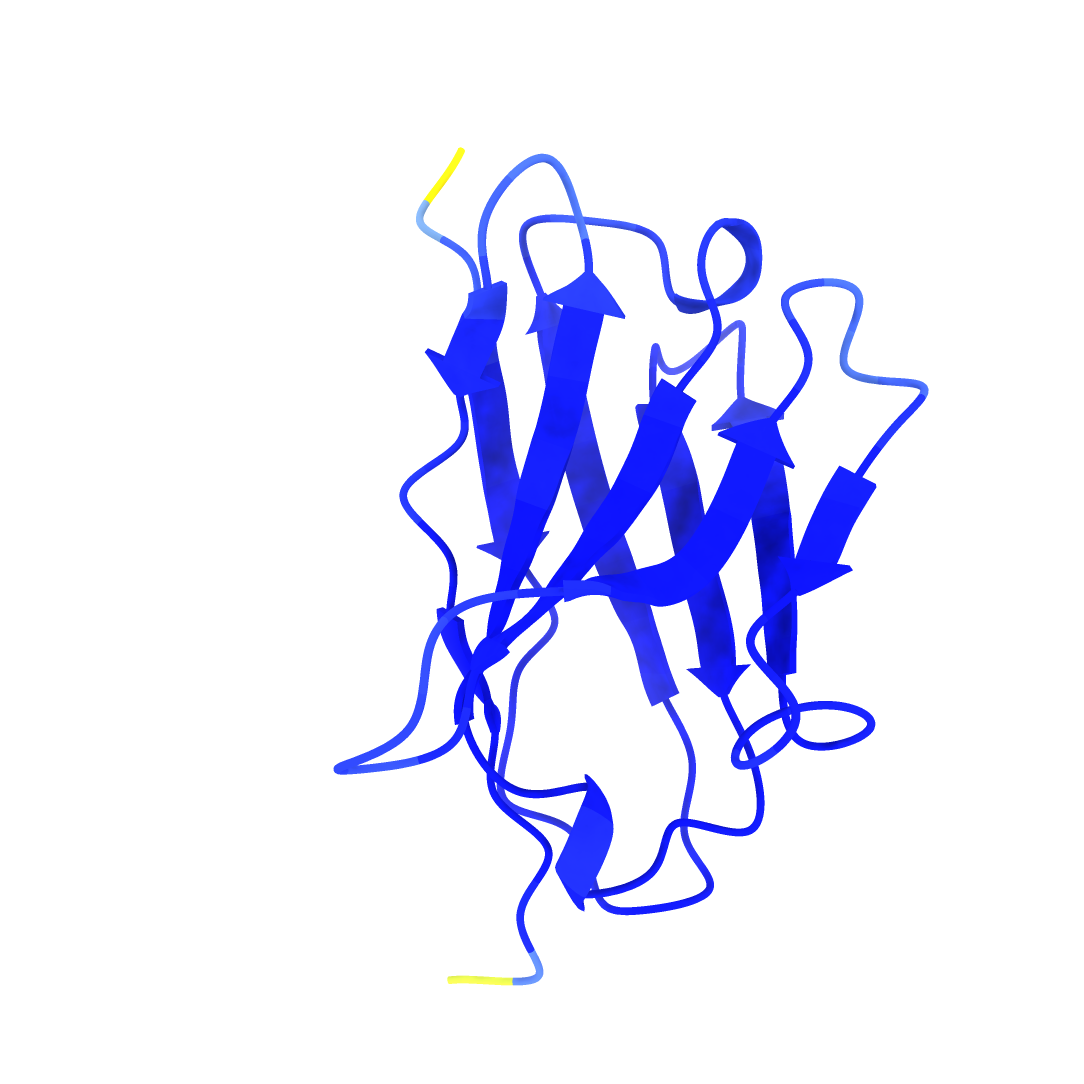
| 
|
| Nb72 nanobody | CDR highlighted |
|---|---|
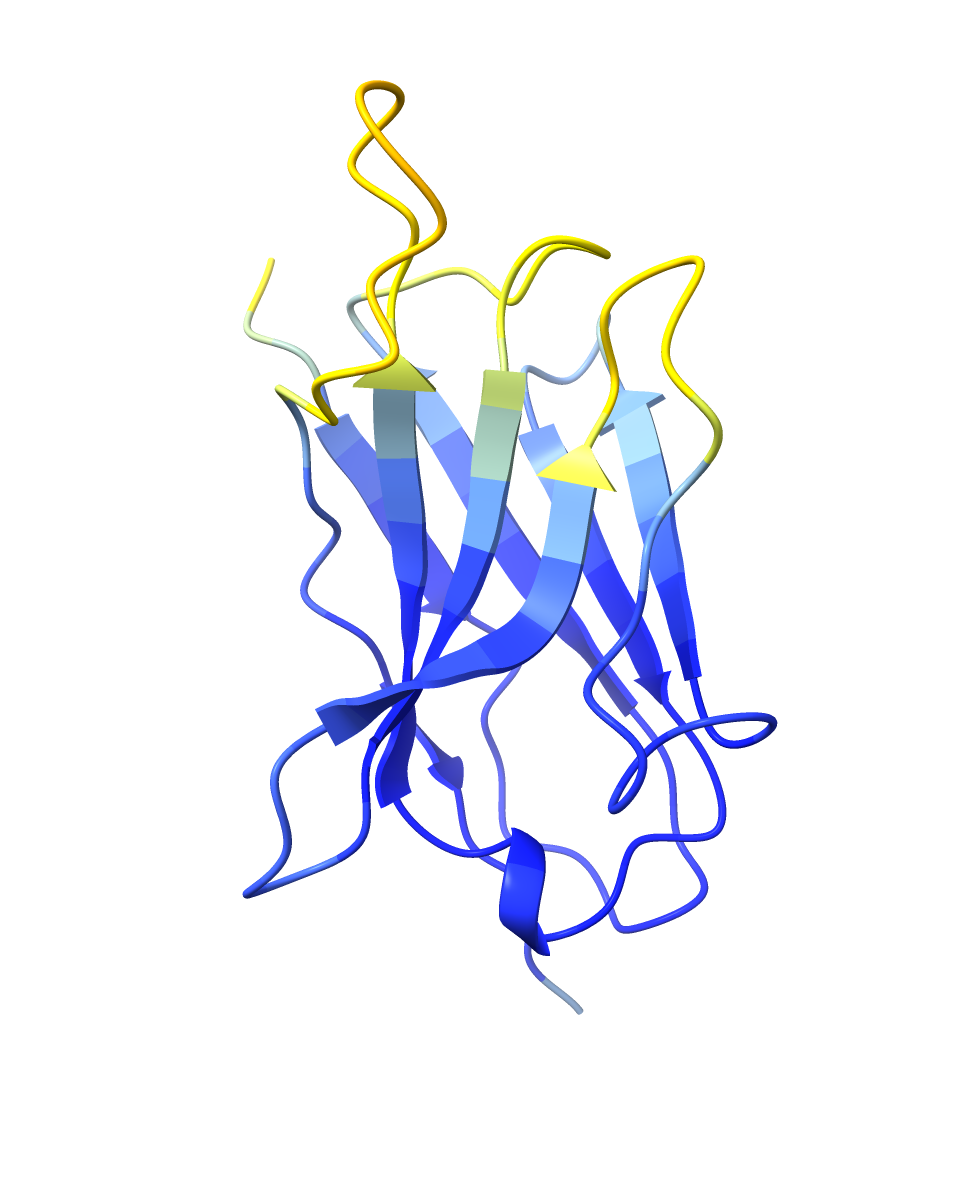
| 
|
Obtain the FRIH structure.
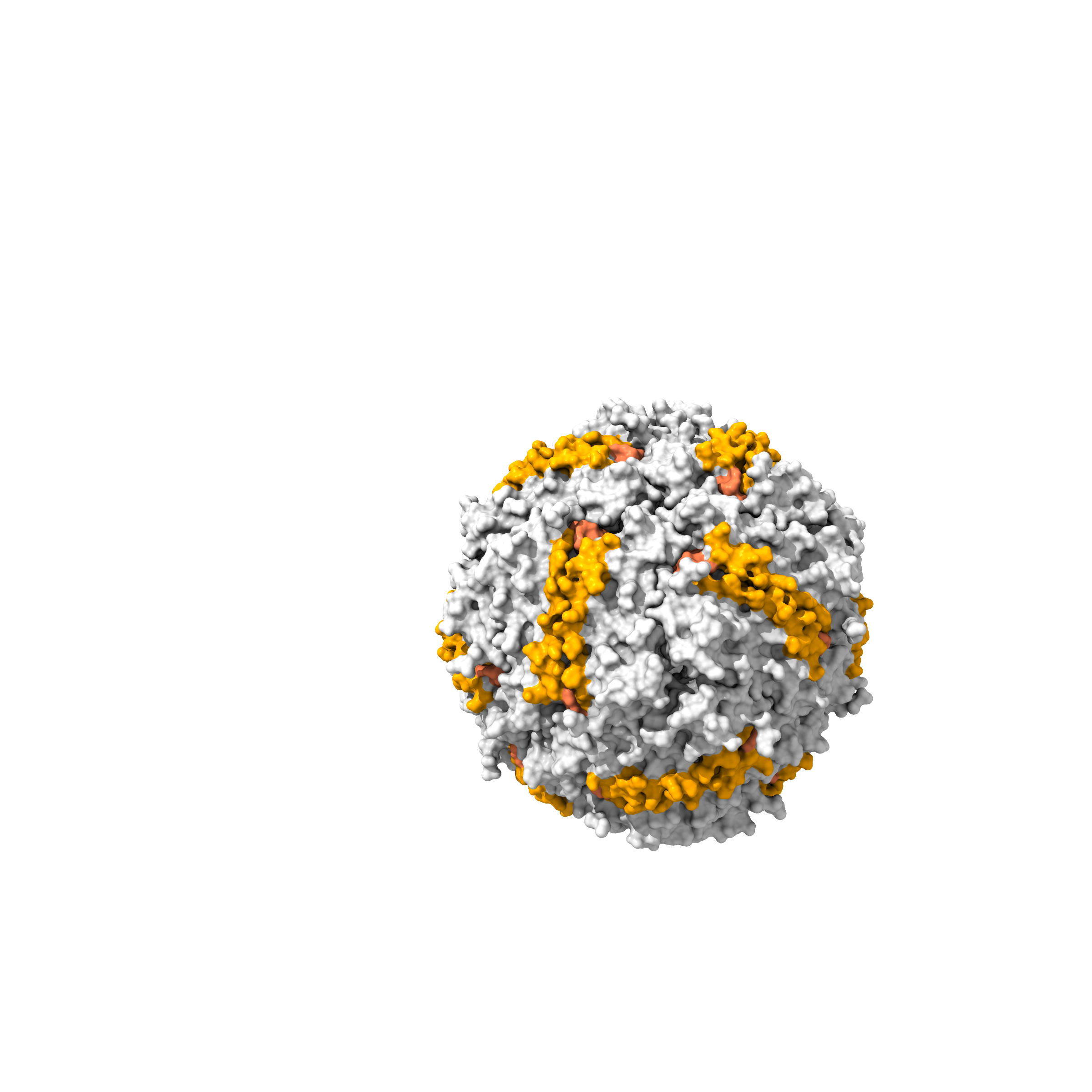
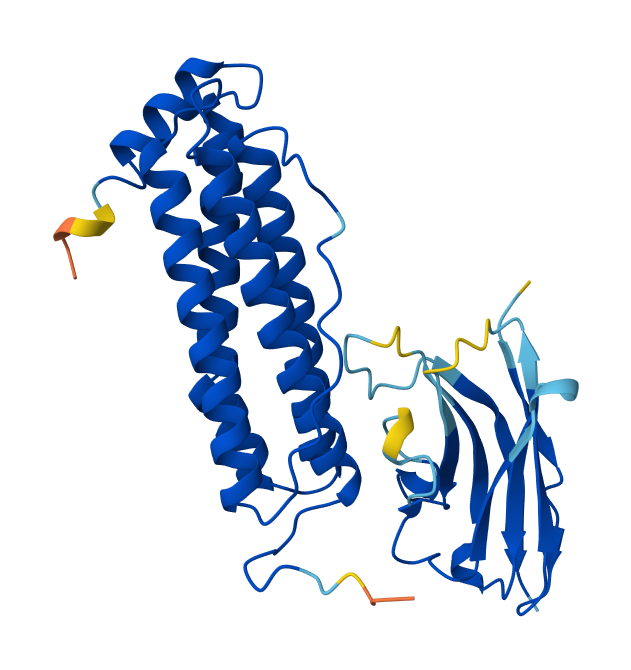
I’ve retrieved the structure of the ferritin heavy chain from the RCSB databank, PDB: 5N27, to locate surface-exposed residues and to use it in docking with the DNA aptamer and the Nb72 nanobody model. The resolution of the 5N27 structure is 1.74 A (Ferraro et al. 2017). Ferritin is a 24-mer cage protein that is hollow inside. Each monomer consists of 183 amino acids.
I’ve visualized the 5N27 structure on ChimeraX and highlighted the surface-exposed residues (A-B and B-C loops) based on the publication by Lawson et. 1991.
The FRIH amino acid sequence, highlighting the A-B and B-C loop residues.
MTTASTSQVRQNYHQDSEAAINRQINLELYASYVYLSMSYYFDRDDVALKNFAKYFLHQSHEEREHAEKLMKLQNQRGGRIFLQDIKKPDCDDWESGLNAMECALHLEKNVNQSLLELHKLATDKNDPHLCDFIETHYLNEQVKAIKELGDHVTNLRKMGAPESGLAEYLFDKHTLGDSDNES
Table showing the boundaries of helices and loops in the FRIH structure.
| Helix | Residue | Amino acid |
|---|---|---|
| A | 13-14 | YHQDSEAAINRQINLELYASYVYLSMSYYF |
| B | 48-77 | ALKNFAKYFLHQSHEEREHAEKLMKLQNQR |
| C | 95-125 | DCDDWESGLNAMECALHLEKNVNQSLLELHK |
| D | 126-159 | LATDKNDPHLCDFIETHYLNEQVKAIKELGDHVT |
| E | 163-174 | KMGAPESGLAEY |
| Loop | Residue | Amino acid |
|---|---|---|
| A-B | 43-47 | DRDDV |
| B-C | 78-94 | GGRIFLQDIKKPDCDDW |
Figures showing the FRIH structure, visualized by ChimeraX, a monomer in a cartoon, and a cage in a surface model. Helices are in blue, the A-B loop is in red, and the B-C loop is in orange.
| FRIH monomer | 24-mer FRIH cage |
|---|---|

| 
|
Docking: Aptamer Seq 20 to FRIH.
Modeling with HADDOCK 2.4
I’ve evaluated the aptamer Seq 20 binding to the FRIH monomer, PDB: 5N27.
The HADDOCK 2.4 modeling uses active residues as input parameters (Honorato et al. 2024 and Honrato et al. 2021).
FRIH.
I've applied FRIH active residues to the solvent-exposed A-B and B-C loops.
Aptamer Seq 20.
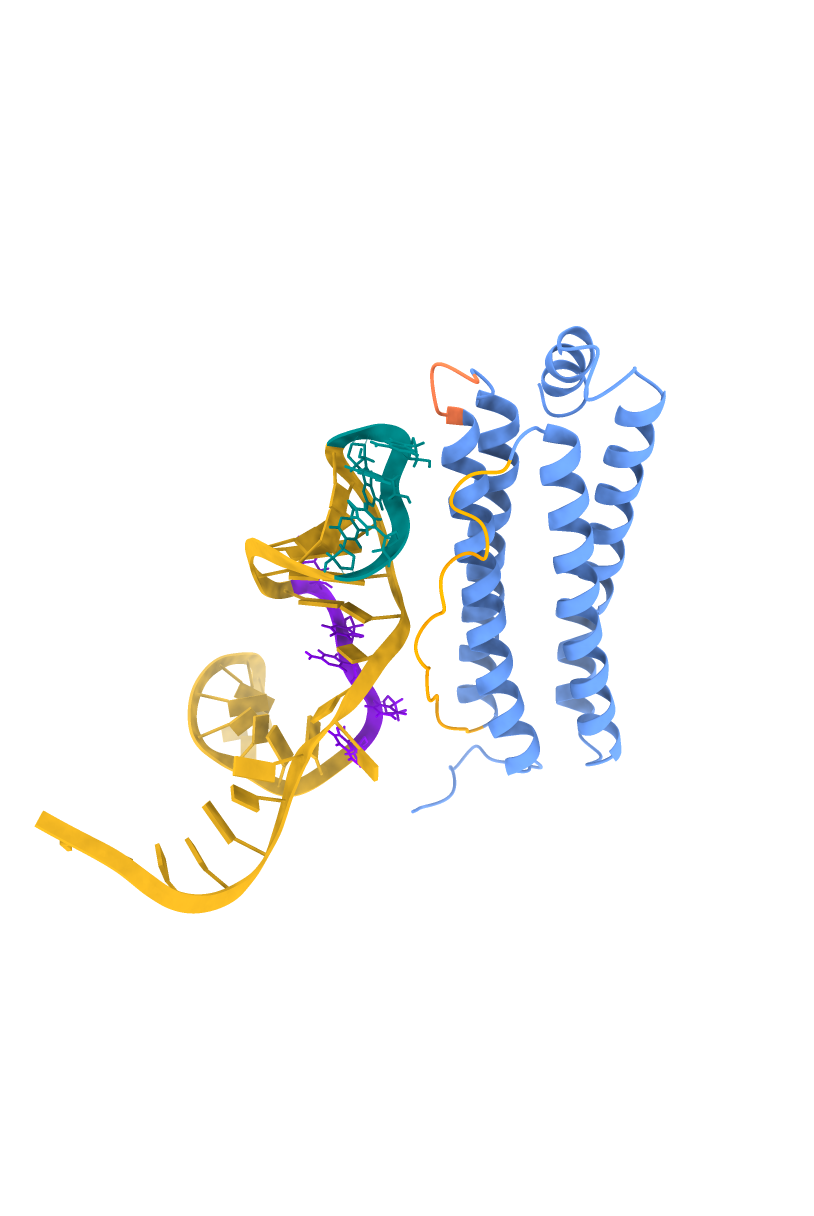
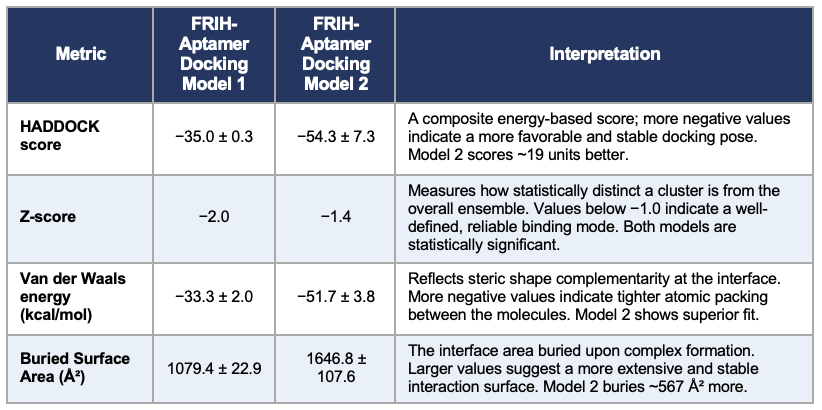
I've applied two approaches: a) considering all predicted high-priority active residues, and 2) testing the hypothesis that a set of active residues may be dominant over other potentially active residues. Model 1 uses the aptamer active residues, primarily the hairpin loop tip residues, 16-20: AAGTC. Model 2 uses both the hairpin loop tip and the stem-loop junction residues, 16-20: AAGTC and 26-30: TTCTG.
The HADDOCK 2.4 scoring indicated that Model 2 is more stable; the score was -54.3. The lower the value, the better. A more stable and extensive interaction surface area was found in the stem-loop junction with a buried surface area of 1646.8 ± 107.6 Ų, having better complementarity, as suggested by the van der Waals energy values, −51.7 ± 3.8 kcal/mol. Both models supported statistically significant and reliable folds.
Figures showing the HADDOCK 2.4 modeling, with aptamer Seq 20 docking to FRIH.
| FRIH-Aptamer Model 1 | FRIH-Aptamer Model 2 |
|---|---|
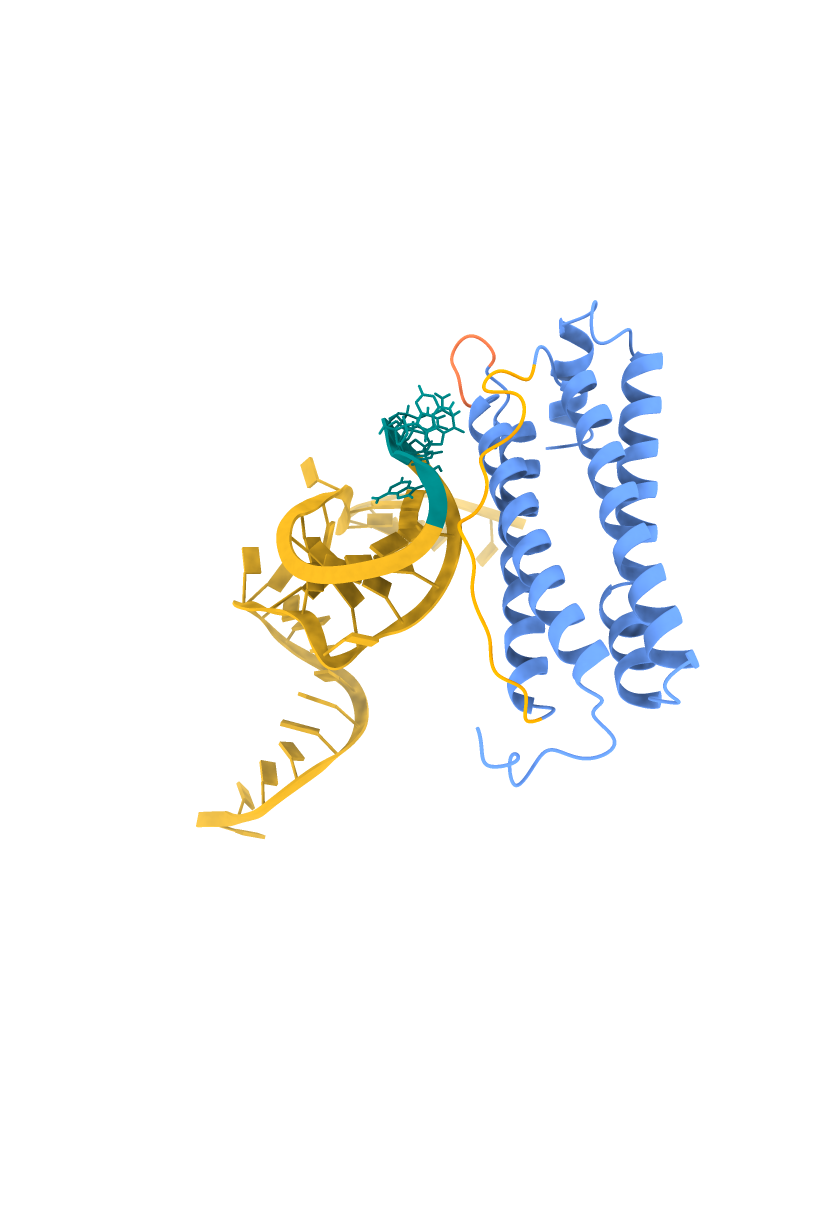
| 
|
FFRIH helices are in blue, surface-exposed loops A-B in coral, B-C in orange. The aptamer Seq 20 is in a schematic model where the high-priority active residues are in a ball-and-stick model. The hairpin loop tip residues (16-20: AAGTC) are in teal, and the stem-loop junction residues (26-30: TTCTG) are in purple.
Summary of the HADDOCK 2.4 modeling comparison.
Docking: Nb72 to FRIH.
Modeling with AlphaFold3
I’ve created a docking model, the Nb72 nanobody to FRIH, using AlphaFold3. The input was the amino acid sequence of the FRIH monomer and the grafted amino acid sequence of the Nb72 nanobody.
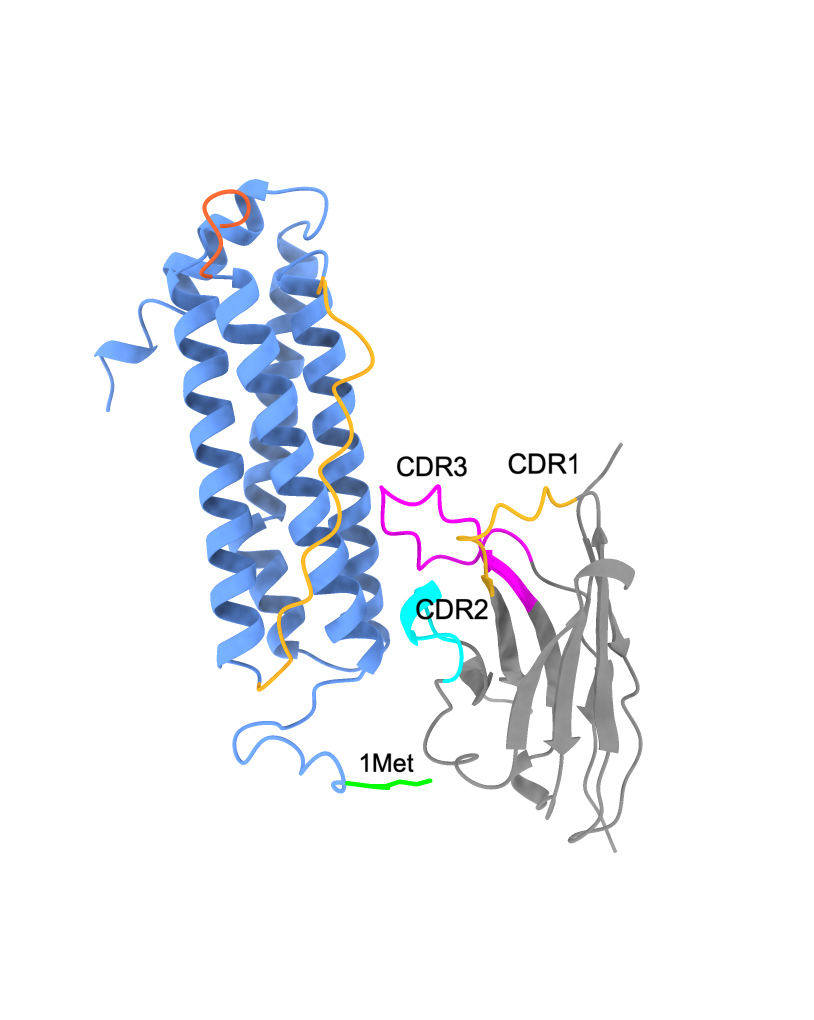
The docking model had a predicted template modeling (pTM) score of 0.77, indicating that the overall predicted fold has high confidence, as the pTM value was above 0.5. The interface predicted template modeling (ipTM) score was 0.6. iPTM measures the accuracy of the predicted relative positions of the subunits within the complex. Higher values, such as 0.8 and greater, are confident, while values of 0.6 and below are low in confidence. The model shows the positioning of Nb72 closer to the N-terminus of FRIH. The CDR3 residues, Ile 107 and Ile 108, are closer to the B-C loop residues, Gln 84, Asp 85, and Ile 86.
AlphaFold3 downloaded image (left), visualization of the same fold in ChimeraX (right). The FRIH monomer is in blue, and the Nb72 nanobody in gray. FRIH loops: A-B in red and B-C in yellow. Nb72 CDR; CDR1 in yellow, CDR2 in cyan, and CDR3 in magenta.
| FRIH-Nb72 Model | Visualized by ChimeraX |
|---|---|
| 
|
MODULE 2
Cell-free protein synthesis construct design for FRIH.
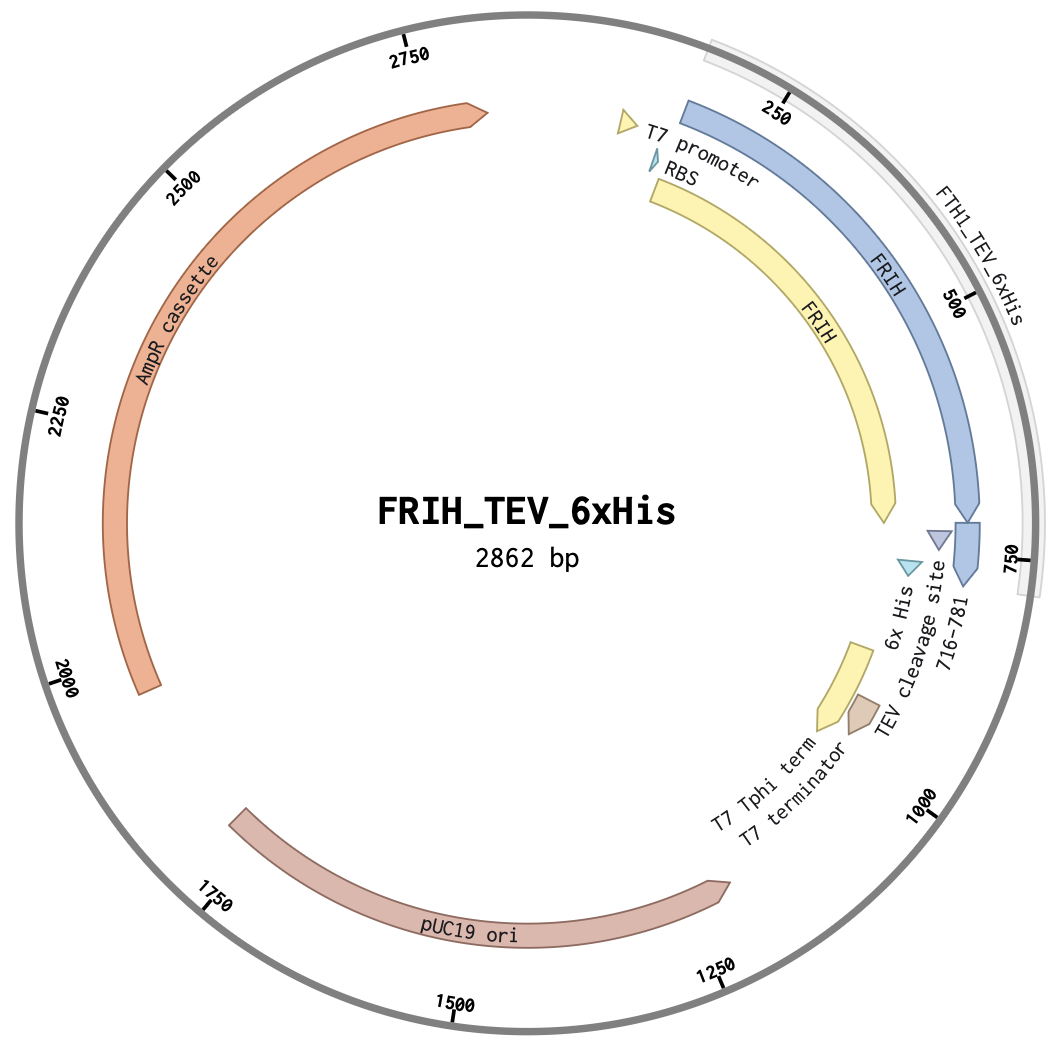
FRIH has been overexpressed from a T7 promoter in an E. coli host and purified (Levi et al. 1987 and Kim et al. 2011). I have proposed to use a cell-free protein synthesis system to express and purify the FRIH protein. For this, I have designed a clonal plasmid construct with T7 promoter, T7 termination, RBS, 6xHis tag, and TEV protease cleavage parts to be cloned at HindIII and SphI sites in pUC19 plasmid by Twist Bio.
The figures shown are the plasmid map of the FRIH T7 expression system and the simulated HindIII and SphI restriction digest, releasing an expected 965 bp fragment, validating the DNA design construct.
Click the link to the Benchling file
Download PDF to view full sequence
| Plasmid map | Restriction digest simulation |
|---|---|
| 
|
MODULE 3
Design of the extended parts, the caged toehold, and the invader strand for the proximity ligation assay (PLA).
PLA is a powerful tool for evaluating the relationship between two distinct epitopes on an antigen. Bound by aptamers or nanobodies, the epitope distance relationships can be revealed by a hybridization event, given that the binders are extended with an oligo arm complementary to each other. When the binders on the antigen are in proximity, the extended arms can interact. A splint connector is added to the ligation reaction that joins the ends, forming a stable dsDNA. A qPCR is generally used as the readout to determine whether the dsDNA is produced.
I’ve designed the extended arms to include a caged toehold; although PLA can be designed differently initially, targeting measuring the epitope distance without the toehold design. FRET probes could be used, for instance, instead of a qPCR readout. Given the design of a caged toehold, which should be validated before PLA, ensuring that branch migration can occur, presumably, it can help to reduce a high background in PLA.
I have applied toehold design principles and generated the DNA sequences listed in the table below (Wang et al. 2015 and Zhang and Winfree 2009). Although computational tools such as the Nupack web server have been widely used for toehold design.

PLA design principle visualized in a diagram.
SUMMARY
Minimally invasive biomarker detection technologies are rapidly advancing toward clinical deployment. The field holds enormous opportunity, but scaling these technologies presents challenges, particularly for large protein biomarkers, whose concentrations in skin interstitial fluid (ISF) are low.
A sandwich-based ELISA, a hybrid detection modality combining 2 distinct binding agents for capturing and reporting, is particularly useful for the detection of large proteins with high specificity. This project focused on the molecular design of such a biosensor for the human ferritin FRIH protein by using computational tools. The biosensor design modeling requires 2 distinct agents to bind in proximity to the FRIH’s surface. Nb72 nanobody, known as a strong binder to FRIH, can be a great capture nanobody, but no DNA aptamer has been described for FRIH.
My approach to designing an FRIH DNA aptamer included a screen from an in silico-generated library of 100 ssDNA 40-mer, and modeling. I advanced one candidate aptamer, Seq 20, based on its binding score and structural compatibility for molecular docking. Application of the Haddock modeling revealed improvement of model predictions when selecting active-residue constraints, a promise that can be improved by an iterative design-build-test cycle.
Using computational modeling, I’ve created a predicted Nb72 model and docking to FRIH with an ipTM, 0.6, a moderate-confidence prediction on the relative positioning of the binder in the complex, where the Nb72 model is positioned near the N-terminus of FRIH. This location can be used as an exclusion criterion for the rational selection of DNA aptamers.
A design of a proximity ligation assay (PLA) was developed, where extended arm sequences are generated and applied in a secondary screen to validate the secondary structure of the DNA aptamers. The proposed PLA can be used as a high-throughput screen with a qPCR readout for evaluating the utility of binders, where they co-bind to distinct parts of the antigen.
SECTION 6: ADDITIONAL INFORMATION
List of References.
Boonkaew, S., P. Teengam, S. Jampasa, S. Rengpipat, W. Siangproh, and O. Chailapakul. 2020. Cost-effective paper-based electrochemical immunosensor using a label-free assay for sensitive detection of ferritin. Analyst.
Ferraro, G., S. Ciambellotti, L. Messori, and A. Merlino. 2017. Cisplatin Binding Sites in Human H-Chain Ferritin. Inorganic Chemistry.
Fredriksson, S., M. Gullberg, J. Jarvius, C. Olsson, K. Pietras, S. M. Gustafsdottir, A. Ostman, and U. Landegren. 2002. Nature Biotechnology.
Han, K., Z. Liang, and N. Zhou. 2010. Design strategies for aptamer-based biosensors. Sensors.
Honorato, R. V., P. J., M.E. Trellet, B. Jiménez-García1, J.J. Schaarschmidt, M. Giulini, V. Reys, P.I. Koukos, J.P.G.L.M. Rodrigues, E. Karaca, G.C.P. van Zundert, J. Roel-Touris, C.W. van Noort, Z. Jandová, A.S.J. Melquiond and A.M.J.J. Bonvin. 2024. “The HADDOCK2.4 web server: A leap forward in integrative modelling of biomolecular complexes” Nature Prot.
Honorato, R. V., P.I. Koukos, B. Jimenez-Garcia, A. Tsaregorodtsev, M. Verlato, A. Giachetti, A. Rosato and A.M.J.J. Bonvin. 2021. “Structural biology in the clouds: The WeNMR-EOSC Ecosystem.” Frontiers Mol. Biosci.
Hu, Y., J. Lin, Y. Wang, S. Wu, J. Wu, H. Lv, X. Ji, S. Muyldermans, Y. Zhang and S. Wang. 2022. Identification of Serum Ferritin-Specific Nanobodies and Development towards a Diagnostic Immunoassay. Biomolecules.
Kim, S-E., K-Y Ahn, J-S Park, K. R. Kim, K. E. Lee, S-S Han, and J. Lee. 2011. Fluorescent Ferritin Nanoparticles and Application to the Aptamer Sensor. Analytical Chemistry.
Klonoff, D. C., D. Ahn, and A. Drincic. 2017. Continuous glucose monitoring: A review of the technology and clinical use. Diabetes Research and Clinical Practice. Diabetes Research and Clinical Practice.
Kubala, M. H., O. Kovtun, K. Alexandrov, and B. M. Collins. 2010. Structural and thermodynamic analysis of the GFP:GFP-nanobody complex. Protein Science.
Lawson, D., P. J. Artymiuk, S. J. Yewdall, J. M. A. Smith, J. C. Livingstone, A. Treffry, A. Luzzago, S. Levi, P. Arosio, G. Cesareni, C. D. Thomas, W. V. Shaw, and P. M. Harrison. 1991. Solving the structure of human H ferritin by genetically engineering intermolecular crystal contacts. Nature.
Levi, S., G. Cesareni, P. Arosio, R. Lorenzetti, M. Soria, M. Sollazzo, A. Albertini and R. Cortese. 1987. Characterization of human ferritin H chain synthesized in Escherichia coli. Gene.
Liang, W. S., B. Beaulieu-Jones, S. Smalley, M. Snyder, L. H. Goetz, and N. J. Schork. 2024. Emerging therapeutic drug monitoring technologies: considerations and opportunities in precision medicine. Frontiers in Pharmacology.
Oharazawa, A., G. Maimaituxun, K. Watanabe, T. Nishiyasu, and N. Fujii. 2024. Metabolome analyses of skin dialysate: insights into skin interstitial fluid biomarkers. Journal of Dermatological Science.
Rodbard, D. 2016. Continuous glucose monitoring: A review of successes, challenges, and opportunities. Diabetes Technology & Therapeutics.
Sprunger, Y., Longo, J., Saeidi, A., Ionescu, A.M. 2025. Bridging Blood and Skin: Biomarker Profiling in Dermal Interstitial Fluid (dISF) for Minimally Invasive Diagnostics. Biosensors.
Vonalioglu, S., M. D. Okur, R. Ustunsoy, T. Ertas, and B. Dinc. 2025. Advances in ferritin biosensors: rapid and cost-effective iron level assessment. Biotech Studies.
Wang, L., L. Fang and S. Liu. 2015. Responsive hairpin DNA aptamer switch to program the strand displacement reaction for the enhanced electrochemical assay of ATP. Analyst.
Xiao, Y., A. A. Lubin, A. J. Heeger, and K. W. Plaxco. 2005. Label-Free Electronic Detection of Thrombin in Blood Serum by Using an Aptamer-Based Sensor. Angew. Chem. Int.
Zhang, D. Y. and E. Winfree. 2009. Control of DNA Strand Displacement Kinetics Using Toehold Exchange. Journal of the American Chemical Society.
The FP7 WeNMR (project# 261572), H2020 West-Life (project# 675858), the EOSC-hub (project# 777536) and the EGI-ACE (project# 101017567) European e-Infrastructure projects are acknowledged for the use of their web portals, which make use of the EGI infrastructure with the dedicated support of CESNET-MCC, INFN-LNL-2, NCG-INGRID-PT, TW-NCHC, IFCA-LCG2, UA-BITP, TR-FC1-ULAKBIM, CSTCLOUD-EGI, IN2P3-CPPM, SURFsara and NIKHEF, and the additional support of the national GRID Initiatives of Belgium, France, Italy, Germany, the Netherlands, Poland, Portugal, Spain, UK, Taiwan and the US Open Science Grid.
One Slide Final Project Description

Final Project Presentation
slide 1
slide 2
slide 3