Week 11 HW: Building Genomes
Part A: The 1,536 Pixel Artwork Canvas | Collective Artwork
Unfortunately, I missed the deadline, I guess I have to become a TA next year (hurray)! ¯_(ツ)_/¯
I thought the sense of community behind the project was really well designed, and I appericated the idea of many people contributing small pieces to one shared bioartwork.
This also made me think about my own project on protein steganography. It could be interesting to create a message-encoding project where people enter their names, messages, or short memories, and we encode them into DNA or protein sequences. Although this would not be as immediately visual as the pixel canvas, it could still be very aligned with HTGAA. One possible version could even involve producing the encoded DNA in bacteria and sharing it with committed listeners. Adrian from the Ottawa node also suggested that it might be beautiful to encode the information into a plant seed or something similar—a living memorial for HTGAA. :')
 Storyboard generated with GPT Image 2.0
Storyboard generated with GPT Image 2.0
Update on artwork
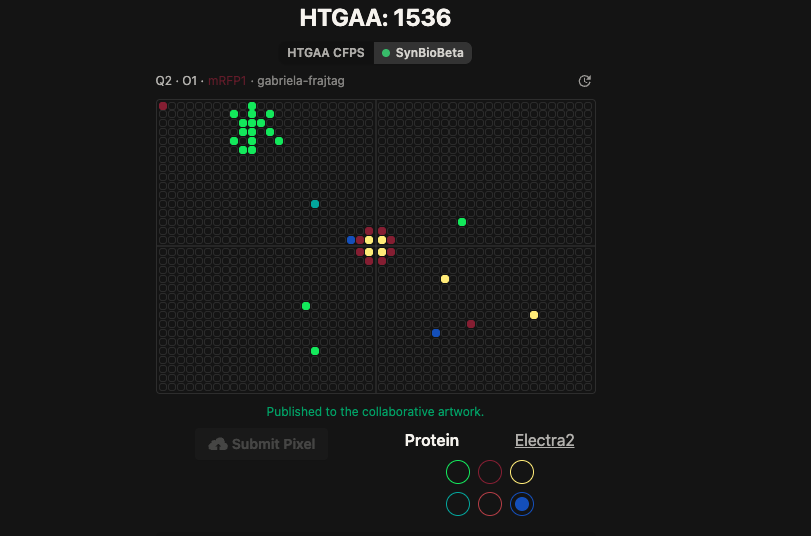 That’s me, the blue one in the middle!
That’s me, the blue one in the middle!
 I took my place in the second version. I’m so happy!
I took my place in the second version. I’m so happy!
Part B: Cell-Free Protein Synthesis | Cell-Free Reagents
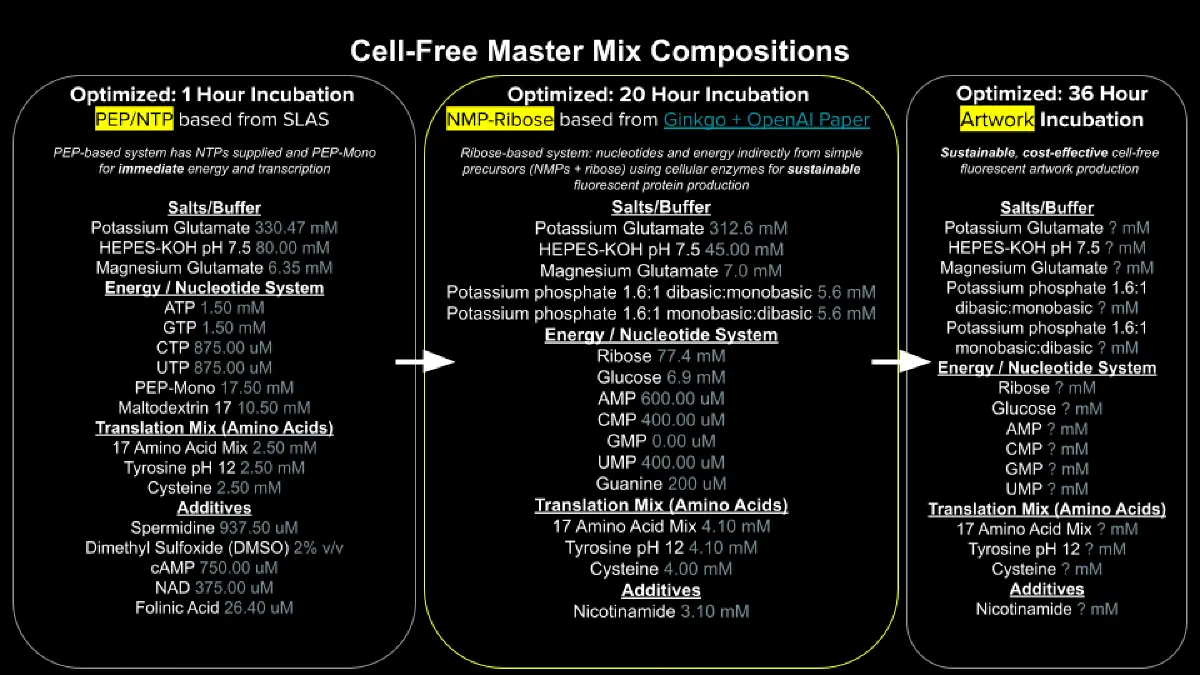
Referencing the cell-free protein synthesis reaction composition (the middle box outlined in yellow on the image above, also listed below), provide a 1-2 sentence description of what each component’s role is in the cell-free reaction.
| Category | Component | Role |
|---|---|---|
| Lysate | BL21(DE3) Star lysate (with T7 RNAP) | Provides the actual machinery doing the work — ribosomes, tRNAs, T7 RNA polymerase, and the metabolic enzymes that recycle ATP and build NTPs. |
| Salts / Buffer | Potassium glutamate | Main potassium source. Glutamate is used instead of chloride because it mimics the inside of E. coli and supports ribosome activity. |
| HEPES-KOH pH 7.5 | Holds the pH near 7.5 as acidic byproducts build up during the reaction. | |
| Magnesium glutamate | Mg²⁺ is essential for RNA polymerase, ribosomes, and almost every step that touches a nucleotide. The level is tuned per lysate batch. | |
| Potassium phosphate (mono- and dibasic) | Supplies the phosphate needed to turn NMPs into NTPs and ADP back into ATP. The mix of mono- and dibasic forms also helps set the starting pH. | |
| Energy / Nucleotide | Ribose | Feeds into PRPP (an activated sugar) used by the salvage pathway to build nucleotides from free bases. |
| Glucose | Cheap energy source. The lysate runs glycolysis on it to regenerate ATP, replacing expensive PEP in long reactions. | |
| AMP, CMP, GMP, UMP | Cheaper monophosphate forms of the nucleotides. Lysate kinases phosphorylate them up to NTPs for transcription. | |
| Guanine | Free purine base that enters the GTP pool through the salvage pathway (see Q3). Cheaper than GMP or GTP. | |
| Translation Mix | 17 AA mix | The 20 standard amino acids minus tyrosine and cysteine, which need separate handling. These are the building blocks for the protein. |
| Tyrosine | Added separately because it doesn’t dissolve well, and because it’s part of the chromophore in most fluorescent proteins — often a limiting reagent for FP yield. | |
| Cysteine | Added separately because it oxidizes easily and forms disulfide bonds, so it’s sensitive to the reaction’s redox state. | |
| Additives | Nicotinamide | Precursor for NAD⁺/NADH. Glycolysis needs NAD⁺ to keep running, which matters when glucose is the energy source. |
| Backfill | Nuclease-free water | Brings everything to the right volume; nuclease-free so the mRNA and DNA template don’t get chewed up. |
Describe the main differences between the 1-hour optimized PEP-NTP master mix and the 20-hour NMP-Ribose-Glucose master mix shown in the Google Slide above.
The PEP-NTP system hands the reaction everything pre-made: NTPs go straight into transcription, and PEP rapidly regenerates ATP from ADP. This produces protein quickly but burns out within an hour as PEP runs out and pyruvate and phosphate build up and acidify the mix — and it’s expensive. The NMP-Ribose-Glucose system instead supplies cheap precursors (NMPs and glucose) and lets the lysate’s own metabolism phosphorylate them up to NTPs and recycle ATP through glycolysis. It starts slower but keeps producing protein for 20+ hours at a fraction of the cost.
How can transcription occur if GMP is not included but Guanine is?
The lysate still has E. coli’s purine salvage pathway intact. An enzyme called XGPRT (Gpt) combines free guanine with PRPP — an activated sugar made from the ribose in the mix — to form GMP. From there, lysate kinases phosphorylate GMP to GDP and then to GTP, which T7 RNA polymerase uses for transcription. So guanine works as a much cheaper drop-in for GMP, with the tradeoff that the conversion is slower — which is fine, even useful, in a long reaction since it avoids burning through the GTP pool too fast.
Part C: Planning the Global Experiment | Cell-Free Master Mix Design
Given the 6 fluorescent proteins we used for our collaborative painting, identify and explain at least one biophysical or functional property of each protein that affects expression or readout in cell-free systems. (Hint: options include maturation time, acid sensitivity, folding, oxygen dependence, etc) (1-2 sentences each)
| Protein | Color | Property affecting CFPS readout |
|---|---|---|
| sfGFP | Green (~510 nm) | Folds fast and matures fast, which is why it’s the standard CFPS reporter. The final maturation step still needs molecular O₂, so sealed wells may become oxygen-limited late in a long run. |
| mRFP1 | Red (~607 nm) | Slow-maturing monomer derived from DsRed; matures >10× faster than DsRed but is still much slower than GFPs, with lower brightness per molecule. A long incubation should help, but a large fraction of protein will still be immature at any given timepoint. |
| mKO2 | Orange (~559 nm) | A faster-maturing version of mKO1, with unusually strong pH stability — meaning it should tolerate the gradual acidification of a long CFPS reaction well. The chromophore contains a cysteine, so it could potentially be sensitive to redox conditions. |
| mTurquoise2 | Cyan (~474 nm) | The brightest cyan FP available (quantum yield ~0.93), but maturation is multi-step and the oxygen-dependent step is rate-limiting. Likely benefits from sustained O₂ supply and stable pH over the 36-hour run. |
| mScarlet-I | Red (~594 nm) | A faster-maturing variant of mScarlet (half-time ~36 min), trading a bit of brightness (QY 0.54) for speed. Practical RFP choice for CFPS since it matures much faster than mRFP1. |
| Electra2 | Blue (ex ~405 / em ~456 nm) | A newer (2022) BFP derived from anemone eqFP611, with brightness similar to mTagBFP2 in mammalian cells. Behavior in E. coli CFPS is less characterized — aggregation has been reported in some contexts, so its folding over 36 h is somewhat uncertain. |
Create a hypothesis for how adjusting one or more reagents in the cell-free mastermix could improve a specific biophysical or functional property you identified above, in order to maximize fluorescence over a 36-hour incubation. Clearly state the protein, the reagent(s), and the expected effect.
Protein: mRFP1 — slow maturation, two oxidative steps, dimmer per molecule than the other RFPs.
Reagent change: Within the 2 µL adjustable volume, add ~20 mM extra HEPES-KOH (pH 7.5) and ~0.5–1 mM extra tyrosine.
Reasoning and expected effect: Over 36 hours, phosphate, pyruvate, and lactate will accumulate and slowly acidify the reaction. The translation and folding machinery is more pH-sensitive than the mRFP1 chromophore itself, so stronger buffering should keep the ribosomes working productively for longer. Tyrosine is one of the three residues that form the chromophore and is supplied separately because of its low solubility — topping it up directly should reduce the chance it becomes rate-limiting in a long run. The expected outcome is brighter red fluorescence at 36 h compared to the baseline composition.