Projects
Final projects:
- Cell-Free Cortisol Biosensor with Gradient Readout

Psychological stress is a pervasive health concern, yet current methods for monitoring stress biomarkers require electronic devices or clinical interpretation. The exploratory question is whether stress-related biomarkers can instead be detected and displayed using purely biological computation deployable outside of labs. This project aims to develop a cell-free biosensor on a skin-proximate fibrous substrate that detects stress through sweat cortisol and translates it into a visible spatial color gradient.
The hypothesis is that cell-free genetic circuits worn on the skin can detect when sweat cortisol exceeds a threshold and display the result as a visible color change, enabling people to perceive their physiological and mental state easily. Key milestones include validating dual-color spatial patterning on a fibrous substrate using cell-free quorum sensing circuits (Aim 1), integrating a cortisol-responsive aptamer-toehold switch circuit and tuning the detection threshold (Aim 2), and deploying to wearables/skin patches and expanding to multi-analyte (Aim 3).
Methods: cell-free protein synthesis, freeze-dried cell free reactions, LuxI/LuxR quorum sensing, RBS strength tuning for threshold control, DNA aptamer coupled to a toehold switch for cortisol detection.
Demonstrate that cell-free quorum sensing circuits can produce a spatial color output on filter paper. A constitutive T7-LuxI sender circuit expresses LuxI (AHL synthase), which enzymatically produces AHL. AHL diffuses outward and activates two receiver constructs: a strong-RBS receiver driving one color (low activation threshold) and a weak-RBS receiver driving another color (high activation threshold). Validation includes characterizing the AHL dose-response for both receivers, confirming sender-receiver communication in cell-free, and testing spatial gradient formation on filter paper. DNA constructs were designed in Benchling and synthesized by Twist Bioscience, derived from validated Addgene plasmids (#193624, #193625, #193626).
Replace the constitutive sender with a cortisol-responsive circuit. Cortisol in sweat binds a DNA aptamer, releasing a trigger strand that activates a toehold switch driving LuxI (AHL synthase) expression. This links the spatial color gradient directly to sweat cortisol concentration. This aim includes designing the aptamer-toehold-LuxI construct, tuning the detection threshold to physiologically relevant cortisol levels, and validating the full cascade from cortisol input to spatial color output in cell-free.
Deploy the system onto a wearable skin patch and expand to multiple sensor zones, each targeting a different sweat biomarker (e.g., cortisol, lactate), each producing a distinct color gradient that together creates a composite body-state readout. Long-term vision: enabling a new category of diagnostic wearable that is disposable, shelf-stable, and readable by the wearer. It addresses a major barrier in personal health monitoring. The broader concept reframes the body’s invisible biochemistry as something expressive, bridging synthetic biology, wearable design, and personal health.
Pardee et al. (2014, Cell) demonstrated that cell-free gene circuits can be freeze-dried onto paper and reactivated by adding water. This foundational work established that synthetic biology diagnostics could operate outside the lab on inexpensive, portable substrates without living cells. However, these systems produce binary yes/no readouts with no ability to distinguish different concentration levels of an analyte.
Basu et al. (2005, Nature) showed that AHL quorum sensing in living E. coli can generate spatial fluorescence patterns through molecular diffusion on agar plates. However, this required living cells and multi-day incubation, limiting deployment outside laboratories.
This project combines the portability of cell-free paper platforms with the spatial computation of AHL diffusion patterning — a combination that has not been previously demonstrated. Existing cell-free biosensors produce binary measurements; by deploying quorum sensing, it enables threshold-based spatial computation with non-binary output. Different RBS strengths create concentration-dependent activation at distinct thresholds, enabling the system to convey how much of an analyte is present. The system also introduces sequential signal processing to cell-free biosensing: rather than a single receptor directly producing output, the signal passes through multiple stages, where each can be independently tuned.
Chronic stress is a growing public health concern linked to cardiovascular disease, immune dysfunction, and mental health disorders, yet most people have no way to monitor their stress levels in daily life. Existing cortisol monitoring devices are either clinical lab tests or expensive electronic wearable sensors. This creates an accessibility barrier where continuous stress monitoring is available only to those with access. A paper-based biosensor that translates cortisol into a visible pattern reduces the cost and complexity of stress monitoring to the level of a disposable sticker. If the multi-analyte vision is realized, the platform could shift the paradigm of personal health monitoring from device-dependent digital readouts to material-based biological one, where the wearable itself performs the sensing, processing, and display.
This project detects a biomarker associated with emotional and physiological state, raising implications around privacy, autonomy and beneficence. The principle of autonomy requires that the wearer has full control over when to use the device and who can see the result. A visible color change on the body could unintentionally disclose stress to others, so the design must ensure the readout is private to the wearer. The system should genuinely help people understand their state, not create new anxiety. If the readout is inaccurate or misinterpreted, the device could cause harm rather than empowerment, turning a wellness tool into a source of stress itself.
To address these concerns, the sensor should be positioned where only the wearer can observe it, and accompanied by clear communication that it indicates cortisol level, not a clinical diagnosis. A key assumption that could be wrong is that sweat cortisol reliably reflects psychological stress, since cortisol also rises during exercise, illness, or normal circadian cycles. An unintended consequence is dual use: if employers or institutions require wearing the device, it shifts from a personal tool to a surveillance tool, violating the autonomy it was designed to support. Alternative approaches include pairing the readout with contextual information (time of day, activity) or framing it strictly as a reflection tool.
The system uses the LuxI/LuxR quorum sensing pathway, a cell-cell communication system natively found in the marine bacterium Vibrio fischeri (Engebrecht et al., 1983, Cell). Three molecular components are needed:
| Component | Role |
|---|---|
| LuxI (AHL synthase) | Enzyme that synthesizes AHL (3OC6-HSL) from substrates in the cell-free lysate |
| LuxR (transcription factor) | Inactive alone; when AHL binds LuxR, the complex becomes an active transcription factor |
| pLux (promoter) | Only transcribed when the LuxR-AHL complex is bound to it |
Signal flow: LuxI produces AHL → AHL diffuses and binds LuxR → LuxR-AHL activates pLux → pLux drives fluorescent reporter
The original system from the Dora Tang lab (Gonzales et al. 2023, ChemSystemsChem) uses three separate plasmids:
| Plasmid | Function |
|---|---|
| #193625 | T7 → LuxR |
| #193624 | pLux → eGFP |
| #193626 | T7 → LuxI |
Their protocol adds LuxR and pLux-eGFP as two separate DNAs to the same cell-free reaction. I simplified this by combining LuxR production and the output reporter onto a single plasmid, so each receiver is self-contained: one DNA instead of two, LuxR-to-reporter ratio can also be tuned with RBS strength.
Each receiver has two transcription units on one backbone:
- T7 → RBS → LuxR → terminator: T7 RNAP in lysate constitutively produces LuxR.
- pLux → RBS → reporter → terminator: stays silent until AHL + LuxR activates pLux.
LuxR is always on; reporter only turns on when AHL is present.
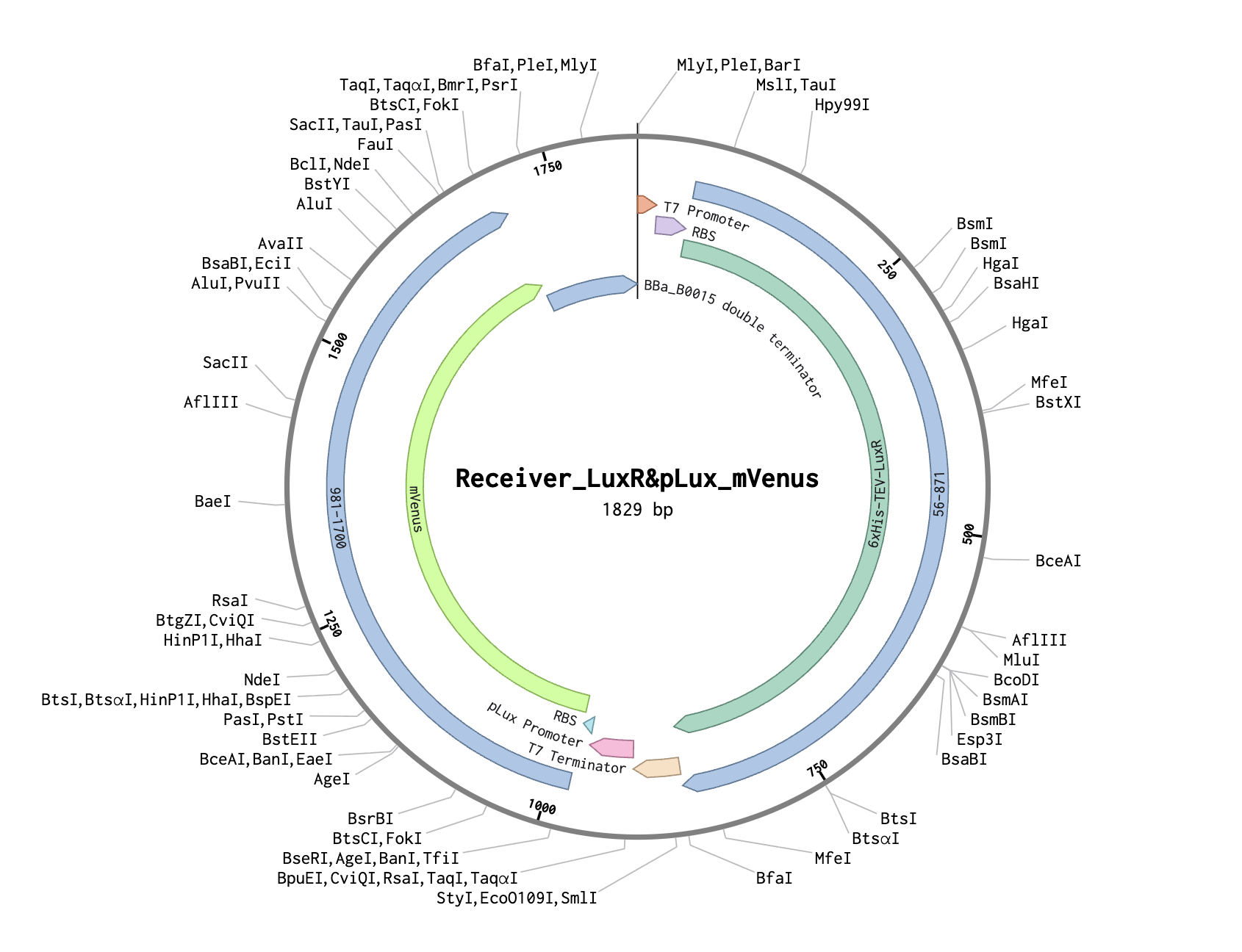
| Part | Description |
|---|---|
| T7 promoter | Constitutive in BL21 DE3 Star lysate |
| Strong RBS (pET RBS) | High translation rate → lots of LuxR protein |
| 6xHis-TEV-LuxR | Transcription factor, from Addgene #193625 |
| T7 terminator | End of first transcription unit |
| pLux promoter | AHL-responsive, from Addgene #193624 |
| RBS (BBa_B0034) | Standard ribosome binding site |
| mVenus | Yellow fluorescent reporter, E. coli codon optimized |
| T7 terminator | End of second transcription unit |
Strong RBS → abundant LuxR → low AHL activates pLux → mVenus turns on far from source.
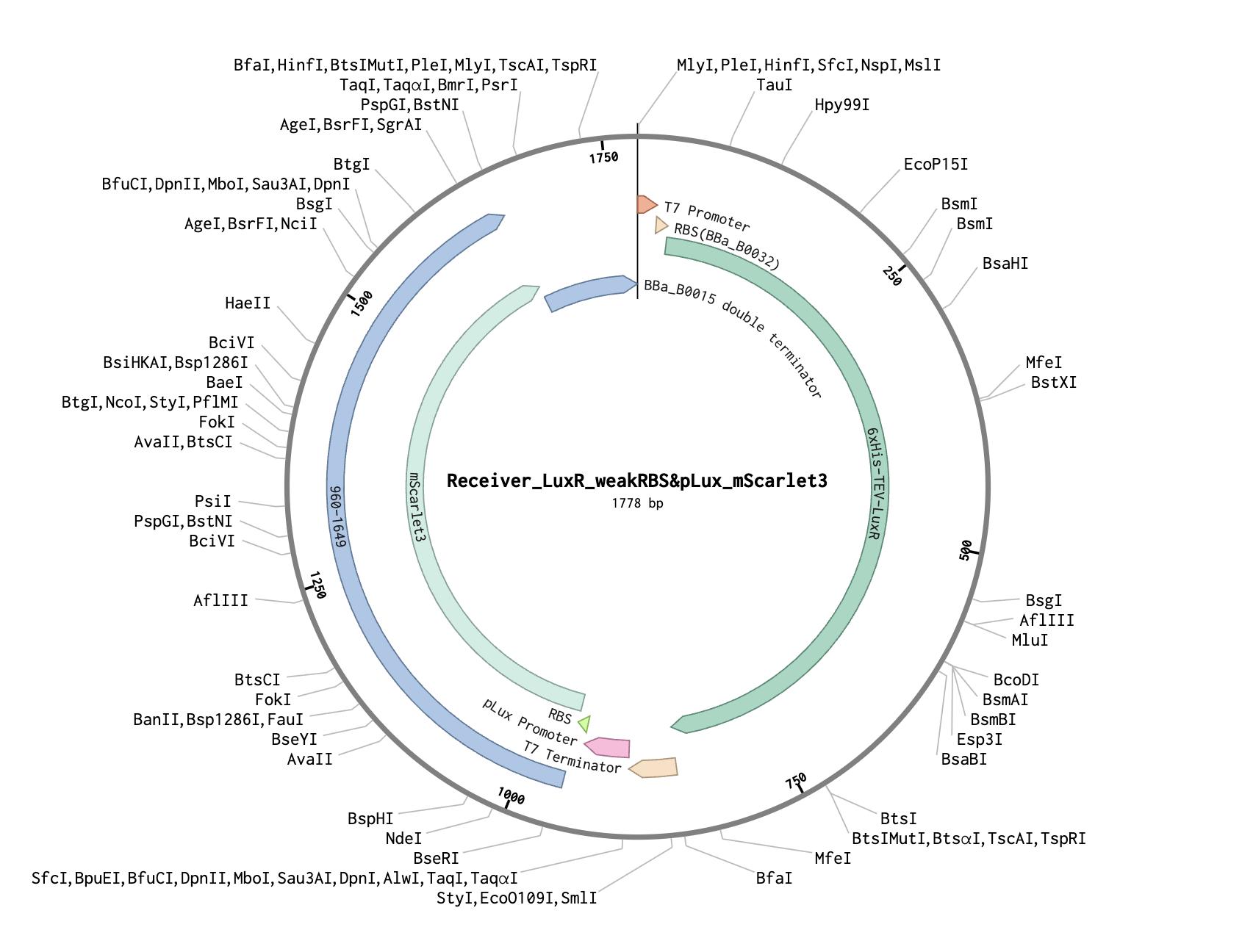
| Part | Description |
|---|---|
| T7 promoter | Constitutive in BL21 DE3 Star lysate |
| Weak RBS (BBa_B0032) | Low translation rate → less LuxR protein |
| 6xHis-TEV-LuxR | Transcription factor, from Addgene #193625 |
| T7 terminator | End of first transcription unit |
| pLux promoter | AHL-responsive, from Addgene #193624 |
| RBS (BBa_B0034) | Standard ribosome binding site |
| mScarlet3 | Red fluorescent reporter, E. coli codon optimized |
| T7 terminator | End of second transcription unit |
Weak RBS → less LuxR → needs high AHL → mScarlet3 only turns on near source.
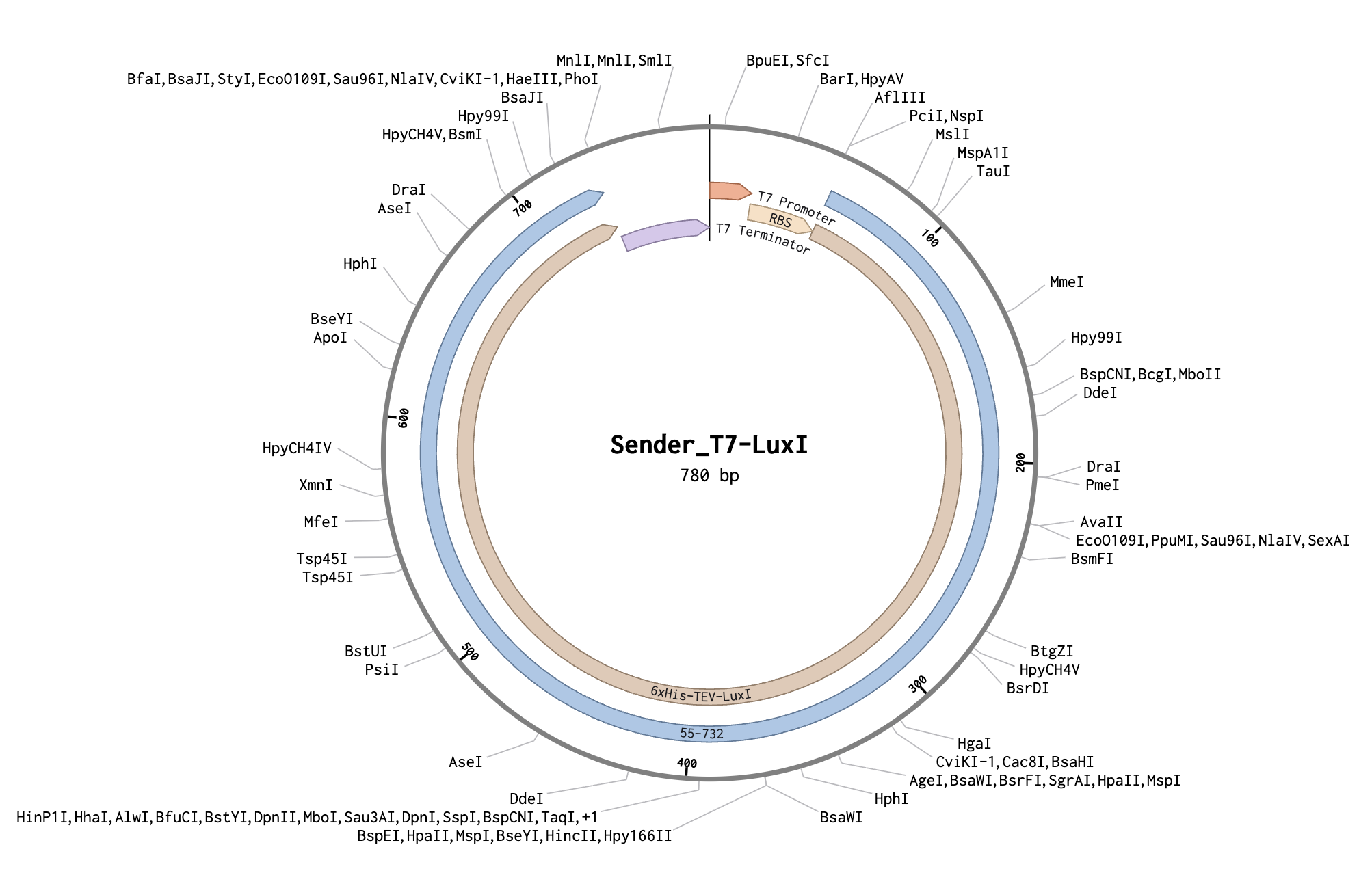
| Part | Description |
|---|---|
| T7 promoter | Constitutive in BL21 DE3 Star lysate |
| RBS (pET RBS) | Standard ribosome binding site |
| 6xHis-TEV-LuxI | AHL synthase, from Addgene #193626 |
| T7 terminator | End transcription |
Constitutively produces LuxI → synthesizes AHL → AHL diffuses outward creating a concentration gradient.
Expected spatial output:
Zone AHL level What turns on Near sender High Both mScarlet3 (red) + mVenus (yellow) Far from sender Low Only mVenus (yellow) Very far None Neither
Before my Twist constructs arrived, I used a constitutive GFP cell-free kit to practice and test on filter paper. The goals were to (1) test whether cell-free reactions produce detectable fluorescence gradients at different DNA concentrations, (2) test whether cell-free reactions work on filter paper without the freeze-dried approach in Pardee’s paper.
 Serial dilution from 50 nM stock
Serial dilution from 50 nM stock
| Tube | Concentration | How to make |
|---|---|---|
| 1 | 50 nM | Original stock |
| 2 | 25 nM | 100 uL of tube 1 + 100 uL water |
| 3 | 12.5 nM | 100 uL of tube 2 + 100 uL water |
| 4 | 6.25 nM | 100 uL of tube 3 + 100 uL water |
| 5 | 3.1 nM | 100 uL of tube 4 + 100 uL water |
| 6 | 1.6 nM | 100 uL of tube 5 + 100 uL water |
| 7 | 0 nM | Nuclease-free water |
For each concentration, I prepared a 60 uL reaction in PCR tubes (kit recipe scaled 3x):
| Component | Volume |
|---|---|
| Reagent Mix | 30 uL |
| Lysate | 18 uL |
| DNA working stock | 6 uL |
| Water | 6 uL |
| Total | 60 uL |
 Pipetting cell-free reactions
Pipetting cell-free reactions
 Reactions ready to go
Reactions ready to go
I tested three layouts on Whatman Grade 1 filter papers:
Radial gradient — dots arranged radially from center outward, highest concentration in the center. Simulates the spatial gradient expected from AHL diffusion.

Linear gradient strips — each concentration deposited as a continuous strip, high to low.

Linear gradient points — each concentration as a single point in a line, high to low.

A key challenge was keeping filter paper wet during incubation. Cell-free reactions require water to function, but liquid on filter paper absorbs and evaporates quickly. I built a simple humidity chamber: petri dish with water on the bottom, filter paper resting on the rim, sealed with the lid. Wet kimwipes in ziplock bags provided additional humidity.
 Humidity chamber design
Humidity chamber design
 Into the warm room
Into the warm room
At 22 hours, faint fluorescence began appearing:
 22 hours — faint but promising
22 hours — faint but promising
At 42 hours, results were clearly visible. The linear gradient points were the most successful because they had no direct contact with water but remained humid. The linear gradient strips also showed clear fluorescence despite partial dilution from contact with water. The radial gradient was barely visible due to insufficient volume per spot.
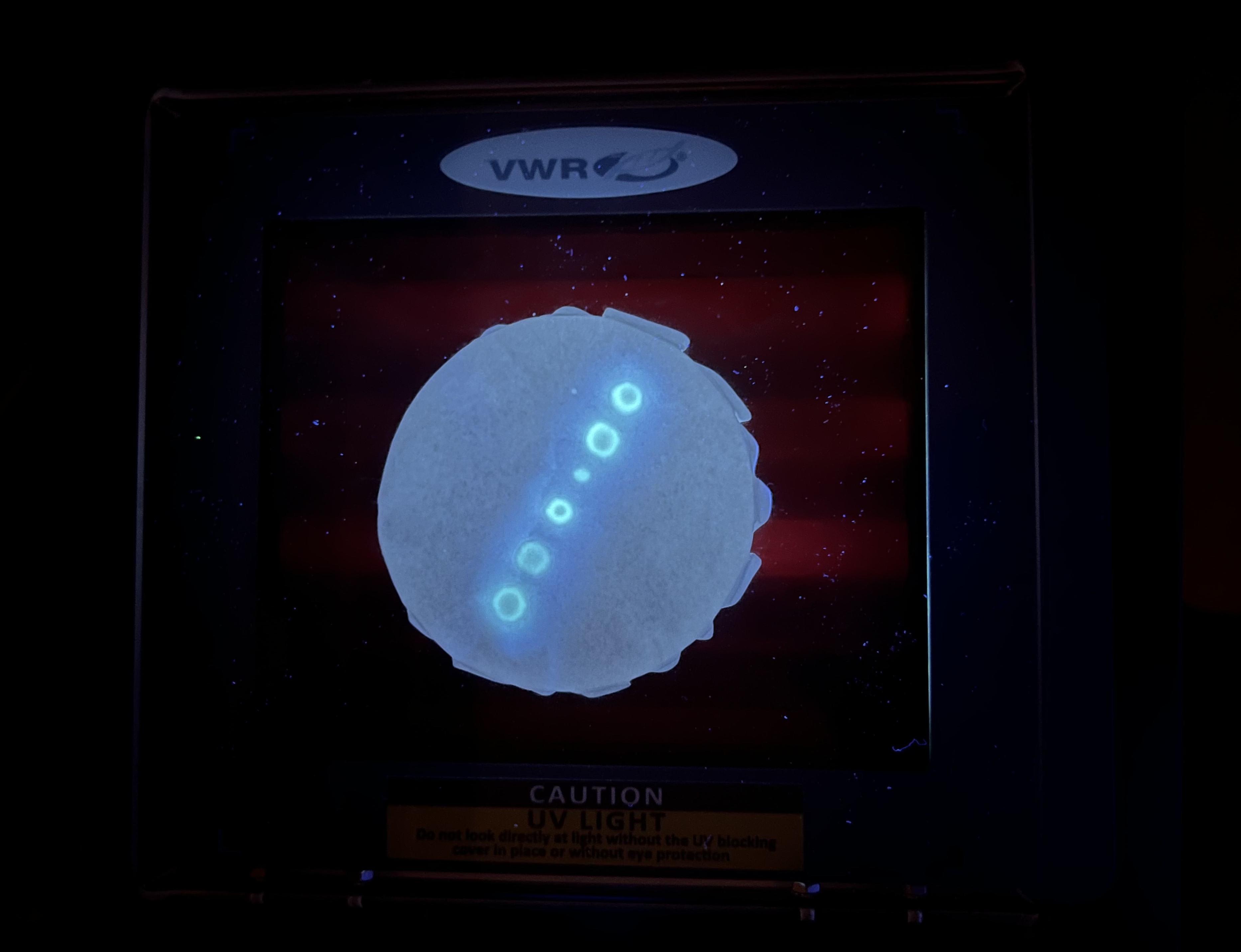
 Successful fluorescent linear spot
Successful fluorescent linear spot
 Diluted linear strips
Diluted linear strips
 All three layouts at 42 hours
All three layouts at 42 hours
The remaining reactions in PCR tubes confirmed GFP expression across all concentrations.
 GFP fluorescence visible across concentration series
GFP fluorescence visible across concentration series
Validated: cell-free reactions can produce visible fluorescent protein on filter paper — a preliminary proof of the core platform for cell-free circuits on a fibrous substrate.
Key takeaways: Paper-based reactions needed significantly longer incubation than tube reactions. Volume per spot is critical. The primary challenge was maintaining humidity without dilution. Protocol adjustments for the actual DNA experiments: larger deposition volumes (15-25 uL) and more careful humidity chamber design.
 The moment of truth
The moment of truth
Of the three ordered constructs, only two were successfully synthesized and delivered: the weak-RBS mScarlet3 receiver (5885 ng) and the constitutive T7-LuxI sender (555 ng). The strong-RBS mVenus receiver failed Twist synthesis. This meant all experiments would use only the high-threshold receiver, and the planned dual-color gradient comparison could not be tested in this round.

DNA resuspension:
| DNA | Total amount | Water added | Final concentration |
|---|---|---|---|
| Receiver (weak RBS, mScarlet3) | 5885 ng | 59 uL | ~100 ng/uL |
| Sender (T7-LuxI) | 555 ng | 22 uL | ~25 ng/uL |
AHL stock preparation: Dissolved 10 mg AHL powder (Sigma K3007, MW 213.23) in 469 uL DMSO → 100 mM main stock. Diluted to 1 mM water-based stock (10 uL stock + 990 uL water), then serial diluted into full-log and half-log working stocks:
 AHL powder dissolving in DMSO
AHL powder dissolving in DMSO
| Tube | Label | How to make |
|---|---|---|
| 1 | AHL-1mM | From 1 mM stock |
| 2 | AHL-100uM | 100 uL of tube 1 + 900 uL water |
| 3 | AHL-10uM | 100 uL of tube 2 + 900 uL water |
| 4 | AHL-1uM | 100 uL of tube 3 + 900 uL water |
| 5 | AHL-100nM | 100 uL of tube 4 + 900 uL water |
| 6 | AHL-300uM | 300 uL of tube 1 + 700 uL water |
| 7 | AHL-30uM | 300 uL of tube 2 + 700 uL water |
| 8 | AHL-3uM | 300 uL of tube 3 + 700 uL water |
| 9 | AHL-300nM | 300 uL of tube 4 + 700 uL water |
| 10 | AHL-30nM | 300 uL of tube 5 + 700 uL water |
 10 working stocks spanning 5 orders of magnitude
10 working stocks spanning 5 orders of magnitude
Purpose: Determine the AHL activation threshold for the weak-RBS mScarlet3 receiver. Expected: Sigmoidal dose-response — low AHL = no fluorescence, high AHL = saturated, inflection = threshold.
Each reaction: 30 uL. All wells identical except AHL concentration.
| Well | Reagent Mix | Lysate | Receiver DNA (100ng/uL) | AHL (3 uL) | Water | Total | Final [AHL] |
|---|---|---|---|---|---|---|---|
| A1 | 15 uL | 9 uL | 3 uL | Tube 1 (1mM) | 0 | 30 uL | 100 uM |
| A2 | 15 uL | 9 uL | 3 uL | Tube 6 (300uM) | 0 | 30 uL | 30 uM |
| A3 | 15 uL | 9 uL | 3 uL | Tube 2 (100uM) | 0 | 30 uL | 10 uM |
| A4 | 15 uL | 9 uL | 3 uL | Tube 7 (30uM) | 0 | 30 uL | 3 uM |
| A5 | 15 uL | 9 uL | 3 uL | Tube 3 (10uM) | 0 | 30 uL | 1 uM |
| A6 | 15 uL | 9 uL | 3 uL | Tube 8 (3uM) | 0 | 30 uL | 300 nM |
| A7 | 15 uL | 9 uL | 3 uL | Tube 4 (1uM) | 0 | 30 uL | 100 nM |
| A8 | 15 uL | 9 uL | 3 uL | Tube 9 (300nM) | 0 | 30 uL | 30 nM |
| A9 | 15 uL | 9 uL | 3 uL | Tube 5 (100nM) | 0 | 30 uL | 10 nM |
| A10 | 15 uL | 9 uL | 3 uL | Tube 10 (30nM) | 0 | 30 uL | 3 nM |
| A11 | 15 uL | 9 uL | 3 uL | none | 3 uL | 30 uL | 0 |
| A12 | 15 uL | 9 uL | 0 | none | 6 uL | 30 uL | no DNA |
Purpose: Test whether LuxI sender produces enough AHL to activate the receiver. Expected: B1 > B2 > B3 > B4 (dose-dependent). B5 confirms receiver works.
| Well | Reagent Mix | Lysate | Receiver DNA (100ng/uL) | Sender DNA (25ng/uL) | AHL | Water | Total | Purpose |
|---|---|---|---|---|---|---|---|---|
| B1 | 15 uL | 9 uL | 3 uL | 3 uL (75 ng) | 0 | 0 | 30 uL | High sender |
| B2 | 15 uL | 9 uL | 3 uL | 2 uL (50 ng) | 0 | 1 uL | 30 uL | Medium sender |
| B3 | 15 uL | 9 uL | 3 uL | 1 uL (25 ng) | 0 | 2 uL | 30 uL | Low sender |
| B4 | 15 uL | 9 uL | 3 uL | 0 | 0 | 3 uL | 30 uL | Negative |
| B5 | 15 uL | 9 uL | 3 uL | 0 | Tube 1 → 3 uL | 0 | 30 uL | Positive |
All 17 reactions loaded into the same 96-well plate — Experiment 1 in row A, Experiment 2 in row B.
 17 wells loaded and ready
17 wells loaded and ready
Plate centrifuged to remove air bubbles before reading.

Plate reader set to 37°C, kinetic reads every 30 minutes (mScarlet3: Ex 569 nm / Em 592 nm).
 12+ hours of kinetic data collection
12+ hours of kinetic data collection
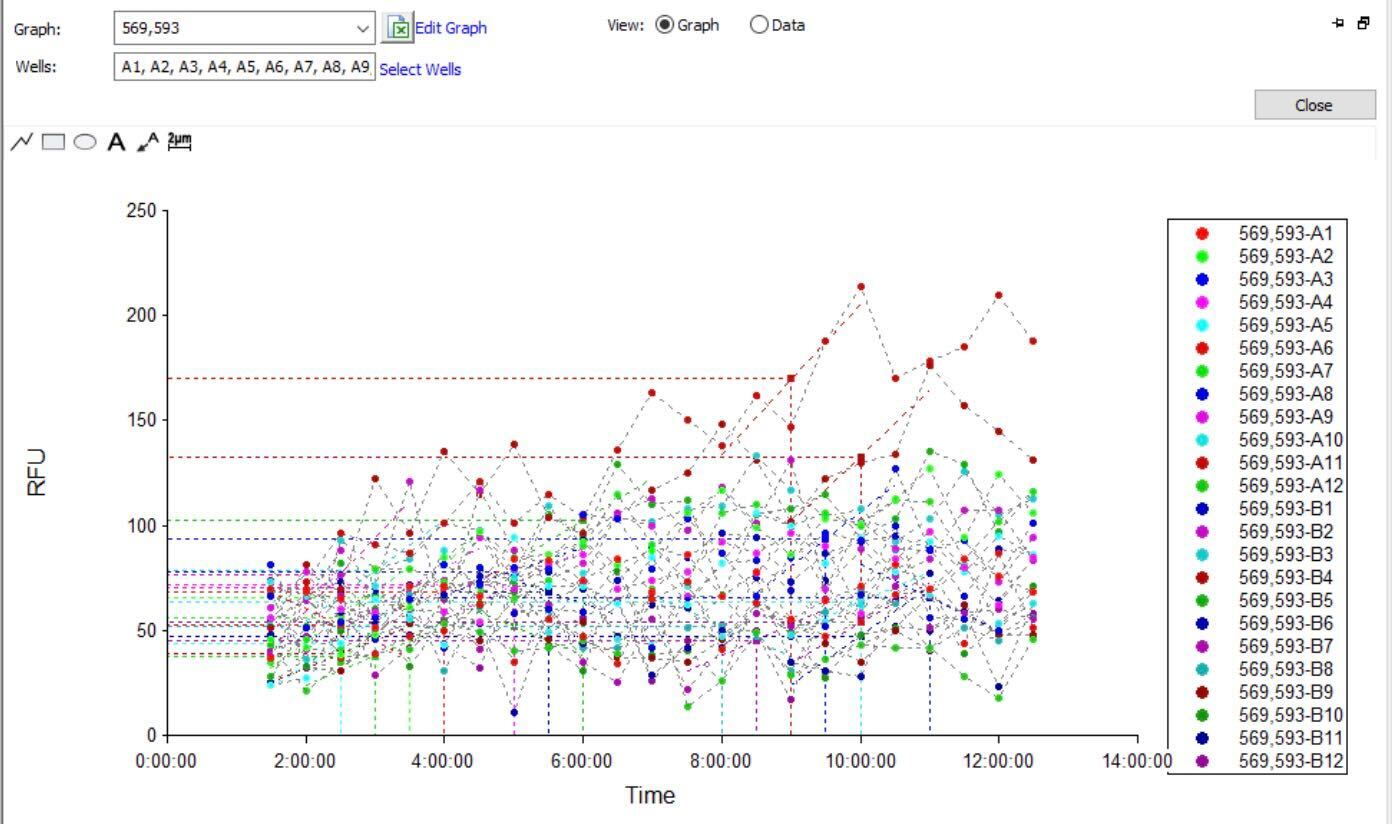 Initial kinetic fluorescence data over ~12 hours
Initial kinetic fluorescence data over ~12 hours
The initial kinetic data showed noisy, low-amplitude signals (30-200 RFU range). We suspected the plastic plate seal was causing optical interference through reflection and scattering. After 12 hours of incubation, we later removed the plate seal and re-read the plate with auto gain enabled, producing much cleaner single-timepoint data:
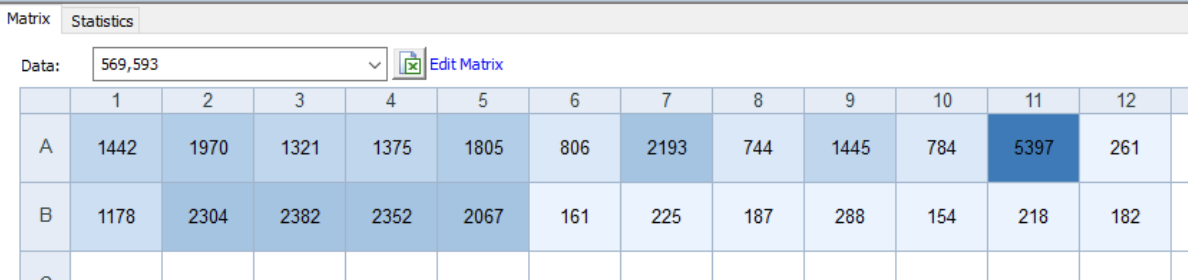 Single read after seal removal with auto gain — signal range now 150-5400 RFU
Single read after seal removal with auto gain — signal range now 150-5400 RFU
Experiment 1 — AHL Dose-Response:
| Well | [AHL] | RFU |
|---|---|---|
| A11 | 0 (no AHL) | 5397 ← highest |
| A7 | 100 nM | 2193 |
| A2 | 30 uM | 1970 |
| A5 | 1 uM | 1805 |
| A9 | 10 nM | 1445 |
| A1 | 100 uM | 1442 |
| A4 | 3 uM | 1375 |
| A3 | 10 uM | 1321 |
| A6 | 300 nM | 806 |
| A10 | 3 nM | 784 |
| A8 | 30 nM | 744 |
| A12 | no DNA | 261 ← baseline |
Experiment 2 — Sender + Receiver:
| Well | Condition | RFU |
|---|---|---|
| B3 | Low sender (25 ng) | 2382 |
| B4 | Negative (no sender) | 2352 |
| B2 | Med sender (50 ng) | 2304 |
| B5 | Positive (AHL) | 2067 |
| B1 | High sender (75 ng) | 1178 ← lowest |
mScarlet3 IS being expressed. A11 = 5397 vs A12 = 261, a 20x difference confirming functional mScarlet3 production from the receiver construct.
However, no AHL dose-response was observed. The no-AHL control (A11) produced the highest signal, and AHL wells showed no sigmoidal trend. Experiment 2 showed a similar inverse pattern: B1 (most sender DNA) had the lowest signal.
Interpretation: T7 read-through + resource competition. The T7 terminator between LuxR and pLux-mScarlet3 does not fully stop T7 RNAP, which reads through and constitutively transcribes mScarlet3 regardless of AHL. This constitutive expression dominates the signal. Adding AHL (with trace DMSO) or sender DNA reduces signal through mild reaction inhibition and resource competition — more components in the reaction means fewer resources available for mScarlet3 production.
What this means: The cell-free system expresses functional fluorescent protein, but the single-plasmid design introduces T7 read-through that masks AHL-responsive behavior. This validates the original Gonzales approach of using separate plasmids for LuxR and the reporter, where T7 read-through from one plasmid cannot reach the reporter on another. Next steps: use a stronger double terminator (BBa_B0015) between the two transcription units, or separate LuxR and the reporter back onto two plasmids.
Two papers prepared for comparison: one with pure AHL at center, one with sender cell-free mix — testing spatial communication on paper.
Receiver mix (one batch, split across both papers):
| Component | Volume |
|---|---|
| Reagent Mix | 50 uL |
| Lysate | 30 uL |
| Receiver DNA (100 ng/uL) | 10 uL (1000 ng) |
| Nuclease-free water | 10 uL |
| Total | 100 uL |
Sender mix (for Paper 2 center):
| Component | Volume |
|---|---|
| Reagent Mix | 30 uL |
| Lysate | 18 uL |
| Sender DNA (25 ng/uL) | 6 uL (150 ng) |
| Nuclease-free water | 6 uL |
| Total | 60 uL |
Receiver spots (15 uL each) deposited radially around a center point on two Whatman Grade 1 filter papers (110 mm).
 Paper 1 — center with pure AHL
Paper 1 — center with pure AHL
 Paper 2 — center with sender cell-free mix
Paper 2 — center with sender cell-free mix
Both papers placed in humidity chambers, incubated at 37°C overnight.
 Humidity chambers sealed and incubating
Humidity chambers sealed and incubating
Results pending; filter paper experiments require extended incubation (24-42 hours based on GFP practice)